–ú–æ—Ä—Ñ–æ–ª–æ–≥–∏—è, —Ñ–∏–∑–∏–æ–ª–æ–≥–∏—è, –±–∏–æ–ª–æ–≥–∏—è
–∏ —Å–∏—Å—Ç–µ–º–∞—Ç–∏–∫–∞ —Ç—Ä–µ–º–∞—Ç–æ–¥.
–ü–æ–ª–æ–≤–æ–∑—Ä–µ–ª—ã–µ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã.
–¢–µ–ª–æ –ø–æ–ª–æ–≤–æ–∑—Ä–µ–ª—ã—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ (–º–∞—Ä–∏—Ç) –≤ –±–æ–ª—å—à–∏–Ω—Å—Ç–≤–µ —Å–ª—É—á–∞–µ–≤ —É–ø–ª–æ—â–µ–Ω–Ω–æ –≤ –¥–æ—Ä—Å–æ-–≤–µ–Ω—Ç—Ä–∞–ª—å–Ω–æ–º –Ω–∞–ø—Ä–∞–≤–ª–µ–Ω–∏–∏, –∏–º–µ–µ—Ç –ª–∏—Å—Ç–æ–≤–∏–¥–Ω—É—é, –ª–∞–Ω—Ü–µ—Ç–æ–≤–∏–¥–Ω—É—é, –∏–Ω–æ–≥–¥–∞ –æ–∫—Ä—É–≥–ª—É—é —Ñ–æ—Ä–º—É. –£–¥–ª–∏–Ω–µ–Ω–Ω–æ–µ —Ç–µ–ª–æ —Å–≤–æ–π—Å—Ç–≤–µ–Ω–Ω–æ —Ç–µ–º —Ç—Ä–µ–º–∞—Ç–æ–¥–∞–º, –∫–æ—Ç–æ—Ä—ã–µ –ª–æ–∫–∞–ª–∏–∑—É—é—Ç—Å—è –≤ —É–∑–∫–∏—Ö –ø—Ä–æ—Ç–æ–∫–∞—Ö (–∂–µ–ª—á–Ω—ã–µ —Ö–æ–¥—ã –ø–µ—á–µ–Ω–∏, –∫—Ä–æ–≤–µ–Ω–æ—Å–Ω—ã–µ —Å–æ—Å—É–¥—ã: —Å–µ–º. Schistosomatidae, Spirorchiidae, Fasciolidae –∏ –¥—Ä.). –¢–∫–∞–Ω–µ–≤—ã–µ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã, –æ–±–∏—Ç–∞—é—â–∏–µ –≤ –æ—Å–æ–±—ã—Ö –∫–∞–ø—Å—É–ª–∞—Ö, –ø—Ä–∏–æ–±—Ä–µ—Ç–∞—é—Ç —à–∞—Ä–æ–≤–∏–¥–Ω—É—é —Ñ–æ—Ä–º—É.
–Ý–∞–∑–º–µ—Ä—ã —Ç—Ä–µ–º–∞—Ç–æ–¥ –∑–∞–≤–∏—Å—è—Ç –æ—Ç —Ä–∞–∑–º–µ—Ä–æ–≤ —Ç–µ–ª–∞ —Ö–æ–∑—è–∏–Ω–∞ –∏ –ª–æ–∫–∞–ª–∏–∑–∞—Ü–∏–∏ –ø–∞—Ä–∞–∑–∏—Ç–∞. –¢–∞–∫, —Ç—Ä–µ–º–∞—Ç–æ–¥—ã, –ø–æ—Å–µ–ª—è—é—â–∏–µ—Å—è –≤ –ø–µ—á–µ–Ω–∏, –∫–∞–∫ –ø—Ä–∞–≤–∏–ª–æ, –∫—Ä—É–ø–Ω–µ–µ —Ä–æ–¥—Å—Ç–≤–µ–Ω–Ω—ã—Ö –∏–º —Ñ–æ—Ä–º, –ª–æ–∫–∞–ª–∏–∑—É—é—â–∏—Ö—Å—è –≤ –∫–∏—à–µ—á–Ω–∏–∫–µ —Ö–æ–∑—è–∏–Ω–∞. –ú–æ–∂–Ω–æ –ø—Ä–æ—Å–ª–µ–¥–∏—Ç—å —è–≤–Ω—É—é —Ç–µ–Ω–¥–µ–Ω—Ü–∏—é –∫ —É–º–µ–Ω—å—à–µ–Ω–∏—é —Ä–∞–∑–º–µ—Ä–æ–≤ –æ—Ç –Ω–∏–∑—à–∏—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ –∫ –≤—ã—Å—à–∏–º; –Ω–∞–∏–±–æ–ª–µ–µ –∫—Ä—É–ø–Ω—ã–µ –≤—Å—Ç—Ä–µ—á–∞—é—Ç—Å—è –≤ —Å–µ–º–µ–π—Å—Ç–≤–∞—Ö Fasciolidae (30-76 –º–º), Cyclocalidae (13-26 –º–º), Echinostomatidae, Paramphistomatidae –∏ –¥—Ä. –í—ã—Å—à–∏–µ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã –∏–º–µ—é—Ç –º–µ–Ω—å—à–∏–µ —Ä–∞–∑–º–µ—Ä—ã.
–£ —Ç—Ä–µ–º–∞—Ç–æ–¥ –∏–º–µ—é—Ç—Å—è –æ—Ä–≥–∞–Ω—ã –ø—Ä–∏–∫—Ä–µ–ø–ª–µ–Ω–∏—è (–ø—Ä–∏—Å–æ—Å–∫–∏), –∫–æ—Ç–æ—Ä—ã–µ –ø—Ä–µ–¥—Å—Ç–∞–≤–ª—è—é—Ç —Å–æ–±–æ–π –º—É—Å–∫—É–ª—å–Ω—ã–µ –≤–∞–ª–∏–∫–∏, —Å–ª–∞–≥–∞—é—â–∏–µ—Å—è –∏–∑ –∫–æ–ª—å—Ü–µ–≤—ã—Ö –∏ —Ä–∞–¥–∏–∞–ª—å–Ω—ã—Ö –º—ã—à–µ—á–Ω—ã—Ö –≤–æ–ª–æ–∫–æ–Ω. –ó–∞ —Å—á–µ—Ç –ø–æ—Å–ª–µ–¥–æ–≤–∞—Ç–µ–ª—å–Ω—ã—Ö —Å–æ–∫—Ä–∞—â–µ–Ω–∏–π –∏ —Ä–∞—Å—Å–ª–∞–±–ª–µ–Ω–∏–π –º—É—Å–∫—É–ª–∞—Ç—É—Ä—ã –ø—Ä–∏—Å–æ—Å–æ–∫ –æ—Å—É—â–µ—Å—Ç–≤–ª—è–µ—Ç—Å—è –ø—Ä–∏–∫—Ä–µ–ø–ª–µ–Ω–∏–µ –ø–∞—Ä–∞–∑–∏—Ç–æ–≤ –∫ —Å—Ç–µ–Ω–∫–∞–º –≤–Ω—É—Ç—Ä–µ–Ω–Ω–∏—Ö –æ—Ä–≥–∞–Ω–æ–≤ —Ö–æ–∑—è–∏–Ω–∞.
–ö–∞–∫ –ø—Ä–∞–≤–∏–ª–æ, —Ç—Ä–µ–º–∞—Ç–æ–¥—ã –∏–º–µ—é—Ç –¥–≤–µ –ø—Ä–∏—Å–æ—Å–∫–∏. –ü–µ—Ä–µ–¥–Ω—è—è –ø—Ä–æ–Ω–∏–∑–∞–Ω–∞ —Ä–æ—Ç–æ–≤—ã–º –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ–º –∏ –Ω–∞–∑—ã–≤–∞–µ—Ç—Å—è —Ä–æ—Ç–æ–≤–æ–π –ø—Ä–∏—Å–æ—Å–∫–æ–π. –í—Ç–æ—Ä–∞—è (–±—Ä—é—à–Ω–∞—è) –ø—Ä–∏—Å–æ—Å–∫–∞ —Å–ª—É–∂–∏—Ç –æ—Ä–≥–∞–Ω–æ–º –ø—Ä–∏–∫—Ä–µ–ø–ª–µ–Ω–∏—è; –æ–Ω–∞ –º–æ–∂–µ—Ç –±—ã—Ç—å —Å–º–µ—â–µ–Ω–∞ –∫ –ø–µ—Ä–µ–¥–Ω–µ–º—É –∫–æ–Ω—Ü—É —Ç–µ–ª–∞ (—Å–µ–º.Fasciolidae, Opisthorchidae), –º–æ–∂–µ—Ç —Ä–∞—Å–ø–æ–ª–∞–≥–∞—Ç—å—Å—è –≤ —Å—Ä–µ–¥–Ω–µ–π –µ–≥–æ —á–∞—Å—Ç–∏ (—Å–µ–º.Plagiorchidae, –∏ –¥—Ä.), –∏–ª–∏ —Å–¥–≤–∏–Ω—É—Ç–∞ –∫ –µ–≥–æ –∑–∞–¥–Ω–µ–º—É –∫–æ–Ω—Ü—É (—Å–µ–º. Paramphistomatidae –∏ –¥—Ä.). –°—Ç–µ–ø–µ–Ω—å —Ä–∞–∑–≤–∏—Ç–∏—è –ø—Ä–∏—Å–æ—Å–æ–∫ –∑–∞–≤–∏—Å–∏—Ç –æ—Ç –ª–æ–∫–∞–ª–∏–∑–∞—Ü–∏–∏ —Ç—Ä–µ–º–∞—Ç–æ–¥. –ù–∞–∏–±–æ–ª—å—à–µ–≥–æ —Ä–∞–∑–≤–∏—Ç–∏—è –ø—Ä–∏—Å–æ—Å–æ–∫ –¥–æ—Å—Ç–∏–≥–∞—é—Ç —É —Ç–µ—Ö –≤–∏–¥–æ–≤, –∫–æ—Ç–æ—Ä—ã–µ –æ–±–∏—Ç–∞—é—Ç –≤ –∫–∏—à–µ—á–Ω–∏–∫–µ —Ö–æ–∑—è–∏–Ω–∞, –æ—Å–æ–±–µ–Ω–Ω–æ –≤ –µ–≥–æ –∑–∞–¥–Ω–µ–º –æ—Ç–¥–µ–ª–µ.
–ü–æ–º–∏–º–æ –ø—Ä–∏—Å–æ—Å–æ–∫, –∏–∑–≤–µ—Å—Ç–Ω—É—é —Ä–æ–ª—å –≤ –ø—Ä–æ—Ü–µ—Å—Å–µ –ø—Ä–∏–∫—Ä–µ–ø–ª–µ–Ω–∏—è —Ç—Ä–µ–º–∞—Ç–æ–¥ –∏–≥—Ä–∞—é—Ç –∏ –∫—É—Ç–∏–∫—É–ª—è—Ä–Ω—ã–µ –æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏—è.
–ü–æ–∫—Ä–æ–≤—ã –∏ –∫–æ–∂–Ω–æ-–º—É—Å–∫—É–ª—å–Ω—ã–π –º–µ—à–æ–∫ —Ç—Ä–µ–º–∞—Ç–æ–¥ –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω—ã —Ü–∏—Ç–æ–ø–ª–∞–∑–º–∞—Ç–∏—á–µ—Å–∫–∏–º —Ç–µ–≥—É–º–µ–Ω—Ç–æ–º. –ü–æ—Å–ª–µ–¥–Ω–∏–π –ø–æ–¥—Ä–∞–∑–¥–µ–ª—è–µ—Ç—Å—è –Ω–∞ –Ω–∞—Ä—É–∂–Ω—ã–π –∏ –≤–Ω—É—Ç—Ä–µ–Ω–Ω–∏–π (–ø–æ–≥—Ä—É–∂–µ–Ω–Ω—ã–π) —É—á–∞—Å—Ç–∫–∏. –í–µ—Å—å –Ω–∞—Ä—É–∂–Ω—ã–π —É—á–∞—Å—Ç–æ–∫ —Ç–µ–≥—É–º–µ–Ω—Ç–∞ –ø—Ä–µ–¥—Å—Ç–∞–≤–ª—è–µ—Ç —Å–æ–±–æ–π –±–µ–∑—ä—è–¥–µ—Ä–Ω—ã–π —Å–∏–Ω—Ü–∏—Ç–∏–π. –û–Ω –æ–≥—Ä–∞–Ω–∏—á–µ–Ω —Å–Ω–∏–∑—É —Ç–æ–Ω–∫–æ–π –±–∞–∑–∞–ª—å–Ω–æ–π –º–µ–º–±—Ä–∞–Ω–æ–π, –∫–æ—Ç–æ—Ä–∞—è –≤–æ –º–Ω–æ–≥–∏—Ö –º–µ—Å—Ç–∞—Ö –ø—Ä–µ—Ä—ã–≤–∞–µ—Ç—Å—è –ø—Ä–æ—Ö–æ–¥—è—â–∏–º–∏ —á–µ—Ä–µ–∑ –Ω–µ–≥–æ –≤—ã—Ä–æ—Å—Ç–∞–º–∏ —Ü–∏—Ç–æ–ø–ª–∞–∑–º—ã, —Å–æ–µ–¥–∏–Ω—è—é—â–µ–π –Ω–∞—Ä—É–∂–Ω—É—é –∏ –≤–Ω—É—Ç—Ä–µ–Ω–Ω—é—é —á–∞—Å—Ç–∏ —Ç–µ–≥—É–º–µ–Ω—Ç–∞. –í –Ω–∞—Ä—É–∂–Ω–æ–º, —Å–∏–Ω—Ü–∏—Ç–∏–∞–ª—å–Ω–æ–º, —É—á–∞—Å—Ç–∫–µ —Ç–µ–≥—É–º–µ–Ω—Ç–∞ —Ä–∞—Å–ø–æ–ª–∞–≥–∞—é—Ç—Å—è –∫—É—Ç–∏–∫—É–ª—è—Ä–Ω—ã–µ —à–∏–ø–∏–∫–∏ –∫–æ–Ω–∏—á–µ—Å–∫–æ–π –∏–ª–∏ –∫—Ä—é—á–∫–æ–≤–∏–¥–Ω–æ–π —Ñ–æ—Ä–º—ã.
–ù–µ–ø–æ—Å—Ä–µ–¥—Å—Ç–≤–µ–Ω–Ω–æ –∑–∞ –±–∞–∑–∞–ª—å–Ω–æ–π –º–µ–º–±—Ä–∞–Ω–æ–π, –æ–≥—Ä–∞–Ω–∏—á–∏–≤–∞—é—â–µ–π —Å–∏–Ω—Ü–∏—Ç–∏–∞–ª—å–Ω—ã–π —É—á–∞—Å—Ç–æ–∫ —Ç–µ–≥—É–º–µ–Ω—Ç–∞ —Å–Ω–∏–∑—É, —Ä–∞—Å–ø–æ–ª–∞–≥–∞—é—Ç—Å—è —Å–ª–æ–∏ –∫–æ–ª—å—Ü–µ–≤—ã—Ö –∏ –ø—Ä–æ–¥–æ–ª—å–Ω—ã—Ö –º—ã—à—Ü, –∫–æ—Ç–æ—Ä—ã–µ –∑–∞–ª–µ–≥–∞—é—Ç –≤ –±–µ—Å—Å—Ç—Ä—É–∫—Ç—É—Ä–Ω–æ–º –º–µ–∂–∫–ª–µ—Ç–æ—á–Ω–æ–º –≤–µ—â–µ—Å—Ç–≤–µ. –í —Ç–æ–ª—â—É –ø–∞—Ä–µ–Ω—Ö–∏–º—ã —Ä–∞—Å–ø–æ–ª–∞–≥–∞—é—Ç—Å—è –ø—É—á–∫–∏ –¥–æ—Ä—Å–æ–≤–µ–Ω—Ç—Ä–∞–ª—å–Ω—ã—Ö –º—ã—à—Ü. –ü–æ–≥—Ä—É–∂–µ–Ω–Ω–∞—è —á–∞—Å—Ç—å —Ç–µ–≥—É–º–µ–Ω—Ç–∞ —Å–æ–¥–µ—Ä–∂–∏—Ç –∫–ª–µ—Ç–æ—á–Ω—ã–µ —ç–ª–µ–º–µ–Ω—Ç—ã —Å —è–¥—Ä–∞–º–∏.
–ú–∞—Ä–∏—Ç—ã —Ç—Ä–µ–º–∞—Ç–æ–¥ –ª–∏—à–µ–Ω—ã –ø–æ–ª–æ—Å—Ç–∏ —Ç–µ–ª–∞, –≤—Å–µ –ø—Ä–æ–º–µ–∂—É—Ç–∫–∏ –º–µ–∂–¥—É –æ—Ä–≥–∞–Ω–∞–º–∏ –∑–∞–ø–æ–ª–Ω–µ–Ω—ã –ø–∞—Ä–µ–Ω—Ö–∏–º–∞—Ç–æ–∑–Ω–æ–π —Ç–∫–∞–Ω—å—é, –≤ —Å–æ—Å—Ç–∞–≤ –∫–æ—Ç–æ—Ä–æ–π –≤—Ö–æ–¥—è—Ç –Ω–µ–¥–∏—Ñ—Ñ–µ—Ä–µ–Ω—Ü–∏—Ä–æ–≤–∞–Ω–Ω—ã–µ –∫–ª–µ—Ç–æ—á–Ω—ã–µ —ç–ª–µ–º–µ–Ω—Ç—ã. –§—É–Ω–∫—Ü–∏–æ–Ω–∞–ª—å–Ω–æ–µ –∑–Ω–∞—á–µ–Ω–∏–µ –ø–∞—Ä–µ–Ω—Ö–∏–º—ã –º–Ω–æ–≥–æ–æ–±—Ä–∞–∑–Ω–æ, –æ–Ω–∞ –∏–≥—Ä–∞–µ—Ç —Ä–æ–ª—å –≤ –ø—Ä–æ—Ü–µ—Å—Å–∞—Ö –æ–±–º–µ–Ω–∞ –≤–µ—â–µ—Å—Ç–≤; –∑–¥–µ—Å—å –Ω–∞–∫–∞–ø–ª–∏–≤–∞–µ—Ç—Å—è –ø–∏—Ç–∞—Ç–µ–ª—å–Ω—ã–µ –≤–µ—â–µ—Å—Ç–≤–∞ (–≥–ª–∏–∫–æ–≥–µ–Ω).
–¢—Ä–µ–º–∞—Ç–æ–¥—ã –∏–º–µ—é—Ç —Ö–æ—Ä–æ—à–æ —Ä–∞–∑–≤–∏—Ç—É—é –ø–∏—â–µ–≤–∞—Ä–∏—Ç–µ–ª—å–Ω—É—é —Å–∏—Å—Ç–µ–º—É. –û–Ω–∞ –Ω–∞—á–∏–Ω–∞–µ—Ç—Å—è —Ä–æ—Ç–æ–≤—ã–º –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ–º, –∫–æ—Ç–æ—Ä–æ–µ –≤–µ–¥–µ—Ç –≤ –º—É—Å–∫—É–ª–∏—Å—Ç—É—é, —à–∞—Ä–æ–≤–∏–¥–Ω—É—é –∏–ª–∏ –æ–≤–∞–ª—å–Ω—É—é –≥–ª–æ—Ç–∫—É. –ó–∞ –≥–ª–æ—Ç–∫–æ–π —Å–ª–µ–¥—É–µ—Ç –ø–∏—â–µ–≤–æ–¥ –∏ –¥–≤—É–≤–µ—Ç–≤–∏—Å—Ç—ã–π –∫–∏—à–µ—á–Ω–∏–∫. –£ –Ω–µ–∫–æ—Ç–æ—Ä—ã—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ (—Å–µ–º.Heterophylidae –∏ –¥—Ä.) —Ä–æ—Ç–æ–≤–æ–µ –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ –æ—Ç–¥–µ–ª–µ–Ω–æ –æ—Ç –≥–ª–æ—Ç–∫–∏ –ø—Ä–µ–¥–≥–ª–æ—Ç–∫–æ–π; –ø–∏—â–µ–≤–æ–¥ –∏–Ω–æ–≥–¥–∞ –æ—Ç—Å—É—Ç—Å—Ç–≤—É–µ—Ç.
–ö–∏—à–µ—á–Ω–∏–∫ —á–∞—â–µ –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω –¥–≤—É–º—è —Å—Ç–≤–æ–ª–∞–º–∏, –∫–æ—Ç–æ—Ä—ã–µ –∏–¥—É—Ç –ø–∞—Ä–∞–ª–ª–µ–ª—å–Ω–æ –±–æ–∫–æ–≤—ã–º –∫—Ä–∞—è–º —Ç–µ–ª–∞ –∏ –∑–∞–∫–∞–Ω—á–∏–≤–∞—é—Ç—Å—è —Å–ª–µ–ø–æ. –ù–µ–ø–µ—Ä–µ–≤–∞—Ä–µ–Ω–Ω—ã–µ –æ—Å—Ç–∞—Ç–∫–∏ –ø–∏—â–∏ –≤—ã–±—Ä–∞—Å—ã–≤–∞—é—Ç—Å—è –Ω–∞—Ä—É–∂—É —á–µ—Ä–µ–∑ —Ä–æ—Ç–æ–≤–æ–µ –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ. –û–¥–Ω–∞–∫–æ —É –Ω–µ–∫–æ—Ç–æ—Ä—ã—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ –∏–º–µ–µ—Ç—Å—è –∏ –∞–Ω–∞–ª—å–Ω–æ–µ –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ. –¢–∞–∫, —É –ø—Ä–µ–¥—Å—Ç–∞–≤–∏—Ç–µ–ª–µ–π —Å–µ–º.Opecoelidae (–ø–∞—Ä–∞–∑–∏—Ç—ã —Ä—ã–±) –∫–æ–Ω—Ü–µ–≤—ã–µ —É—á–∞—Å—Ç–∫–∏ –∫–∏—à–µ—á–Ω—ã—Ö –≤–µ—Ç–≤–µ–π —Å–ª–∏–≤–∞—é—Ç—Å—è –∏ –æ—Ç–∫—Ä—ã–≤–∞—é—Ç—Å—è –Ω–∞—Ä—É–∂—É –æ—Å–æ–±—ã–º –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ–º; —É –¥—Ä—É–≥–∏—Ö –≤–∏–¥–æ–≤ (—Ä.Diploproctoidaeum) –∫–∞–∂–¥—ã–π –∏–∑ –∫–∏—à–µ—á–Ω—ã—Ö —Å—Ç–≤–æ–ª–æ–≤ –æ—Ç–∫—Ä—ã–≤–∞–µ—Ç—Å—è –Ω–∞—Ä—É–∂—É —Å–∞–º–æ—Å—Ç–æ—è—Ç–µ–ª—å–Ω—ã–º –∞–Ω–∞–ª—å–Ω—ã–º –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ–º; —É –ø—Ä–µ–¥—Å—Ç–∞–≤–∏—Ç–µ–ª–µ–π —Å–µ–º.Echinostomatidae –∑–∞–¥–Ω–∏–π –æ—Ç–¥–µ–ª –∫–∏—à–µ—á–Ω—ã—Ö —Å—Ç–≤–æ–ª–æ–≤ —Å–ª–∏–≤–∞–µ—Ç—Å—è —Å —ç–∫—Å–∫—Ä–µ—Ç–æ—Ä–Ω—ã–º –ø—É–∑—ã—Ä–µ–º.
–í—ã–¥–µ–ª–∏—Ç–µ–ª—å–Ω–∞—è —Å–∏—Å—Ç–µ–º–∞ —É –º–∞—Ä–∏—Ç—ã —Å–æ—Å—Ç–æ–∏—Ç –∏–∑ –º–µ—Ä—Ü–∞—Ç–µ–ª—å–Ω—ã—Ö –∫–ª–µ—Ç–æ–∫, —Å–æ–±–∏—Ä–∞—Ç–µ–ª—å–Ω—ã—Ö –∫–∞–Ω–∞–ª—å—Ü–µ–≤ –∏ –º–æ—á–µ–≤–æ–≥–æ (—ç–∫—Å–∫—Ä–µ—Ç–æ—Ä–Ω–æ–≥–æ) –ø—É–∑—ã—Ä—è. –ú–µ—Ä—Ü–∞—Ç–µ–ª—å–Ω—ã–µ –∫–ª–µ—Ç–∫–∏ —É –Ω–µ–∫–æ—Ç–æ—Ä—ã—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ –ª–∏—à–µ–Ω—ã —è–¥—Ä–∞, —á—Ç–æ —Å–ª–µ–¥—É–µ—Ç —Ä–∞—Å—Å–º–∞—Ç—Ä–∏–≤–∞—Ç—å –∫–∞–∫ –ø—Ä–æ—è–≤–ª–µ–Ω–∏–µ —Å–ø–µ—Ü–∏–∞–ª–∏–∑–∞—Ü–∏–∏. –°–æ–±–∏—Ä–∞—Ç–µ–ª—å–Ω—ã–µ –∫–∞–Ω–∞–ª—å—Ü—ã –ø—Ä–µ–¥—Å—Ç–∞–≤–ª—è—é—Ç —Å–æ–±–æ–π –º–Ω–æ–≥–æ–∫–ª–µ—Ç–æ—á–Ω—ã–µ –æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏—è —Å —Å–∏–Ω—Ü–∏—Ç–∏–∞–ª—å–Ω—ã–º–∏ —Å—Ç–µ–Ω–∫–∞–º–∏. –í—Å—è —Å–∏—Å—Ç–µ–º–∞ —ç–∫—Å–∫—Ä–µ—Ç–æ—Ä–Ω—ã—Ö –∫–∞–Ω–∞–ª—å—Ü–µ–≤ –ª–∏—à–µ–Ω–∞ –º—É—Å–∫—É–ª—å–Ω—ã—Ö –≤–æ–ª–æ–∫–æ–Ω. –ú–æ—á–µ–≤–æ–π –ø—É–∑—ã—Ä—å –æ—Ç–∫—Ä—ã–≤–∞–µ—Ç—Å—è –Ω–∞—Ä—É–∂—É –Ω–µ–ø–∞—Ä–Ω—ã–º –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ–º –Ω–∞ –∑–∞–¥–Ω–µ–º –∫–æ–Ω—Ü–µ —Ç–µ–ª–∞, –∫–æ–Ω—Ü–µ–≤–æ–π –æ—Ç–¥–µ–ª –ø—É–∑—ã—Ä—è –Ω–µ—Ä–µ–¥–∫–æ —Å–Ω–∞–±–∂–µ–Ω —Ö–æ—Ä–æ—à–æ —Ä–∞–∑–≤–∏—Ç—ã–º —Å—Ñ–∏–Ω–∫—Ç–µ—Ä–æ–º.
–¶–µ–Ω—Ç—Ä–∞–ª—å–Ω–∞—è —á–∞—Å—Ç—å –Ω–µ—Ä–≤–Ω–æ–π —Å–∏—Å—Ç–µ–º—ã –º–∞—Ä–∏—Ç—ã –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω–∞ –ø–µ—Ä–µ–¥–Ω–∏–º –≥–∞–Ω–≥–ª–∏–µ–º, —Ä–∞—Å–ø–æ–ª–æ–∂–µ–Ω–Ω—ã–º –≤ –ø–µ—Ä–µ–¥–Ω–µ–π —Ç—Ä–µ—Ç–∏ —Ç–µ–ª–∞, —á–∞—Å—Ç–æ –Ω–∞ —É—Ä–æ–≤–Ω–µ –≥–ª–æ—Ç–∫–∏. –ö –ø–µ—Ä–µ–¥–Ω–µ–º—É –∫–æ–Ω—Ü—É —Ç–µ–ª–∞ –æ—Ç –Ω–µ–≥–æ –æ—Ç—Ö–æ–¥—è—Ç —Ç—Ä–∏ –ø–∞—Ä—ã –Ω–µ—Ä–≤–Ω—ã—Ö —Å—Ç–≤–æ–ª–æ–≤, –∫–æ–Ω—Ü–µ–≤—ã–µ —É—á–∞—Å—Ç–∫–∏ –∫–æ—Ç–æ—Ä—ã—Ö —Ä–∞–∑–≤–µ—Ç–≤–ª—è—é—Ç—Å—è –Ω–∞ –º–Ω–æ–∂–µ—Å—Ç–≤–æ –Ω–µ—Ä–≤–Ω—ã—Ö –≤–µ—Ç–æ—á–µ–∫, –∑–∞—Ö–æ–¥—è—â–∏—Ö –≤ —Ç–µ–≥—É–º–µ–Ω—Ç. –ù–∞–∑–∞–¥ –æ—Ç –º–æ–∑–≥–æ–≤–æ–≥–æ –≥–∞–Ω–≥–ª–∏—è –æ—Ç—Ö–æ–¥—è—Ç –¥–≤–∞ –≤–µ–Ω—Ç—Ä–∞–ª—å–Ω—ã—Ö, –¥–≤–∞ –ª–∞—Ç–µ—Ä–∞–ª—å–Ω—ã—Ö –∏ –¥–≤–∞ –¥–æ—Ä—Å–∞–ª—å–Ω—ã—Ö –Ω–µ—Ä–≤–Ω—ã—Ö —Å—Ç–≤–æ–ª–∞, –¥–æ—Å—Ç–∏–≥–∞—é—â–∏—Ö –∑–∞–¥–Ω–µ–≥–æ –∫–æ–Ω—Ü–∞ —Ç–µ–ª–∞, –≥–¥–µ –∫–∞–∂–¥–∞—è –ø–∞—Ä–∞ —Å—Ç–≤–æ–ª–æ–≤ —Å–ª–∏–≤–∞–µ—Ç—Å—è, –æ–±—Ä–∞–∑—É—è –∞—Ä–∫—É. –ü—Ä–æ–¥–æ–ª—å–Ω—ã–µ —Å—Ç–≤–æ–ª—ã —Å–æ–µ–¥–∏–Ω–µ–Ω—ã –º–µ–∂–¥—É —Å–æ–±–æ–π –ø–æ–ø–µ—Ä–µ—á–Ω—ã–º–∏ –∫–æ–º–∏—Å—Å—É—Ä–∞–º–∏, —á–∞—Å—Ç–æ –∏–º–µ—é—â–∏–º–∏ –≤–∏–¥ –ø–æ–ª—É–∫–æ–ª–µ—Ü –∏–ª–∏ –∫–æ–ª–µ—Ü, –æ–ø–æ—è—Å—ã–≤–∞—é—â–∏—Ö —Ç–µ–ª–æ —á–µ—Ä–≤—è.
–ú–∞—Ä–∏—Ç—ã –ø–æ—á—Ç–∏ –ª–∏—à–µ–Ω—ã –æ—Ä–≥–∞–Ω–æ–≤ —á—É–≤—Å—Ç–≤, –ª–∏—à—å —É –Ω–µ–∫–æ—Ç–æ—Ä—ã—Ö —Ä–æ–¥–æ–≤ (Dicrocaelium) –∏–º–µ—é—Ç—Å—è —Å–µ–Ω—Å–∏–ª–ª—ã –Ω–∞ —Ä–æ—Ç–æ–≤–æ–π –ø—Ä–∏—Å–æ—Å–∫–µ. –ò–Ω–æ–≥–¥–∞ —Å–æ—Ö—Ä–∞–Ω—è—é—Ç—Å—è —Ä—É–¥–∏–º–µ–Ω—Ç—ã –≥–ª–∞–∑ —Ü–µ—Ä–∫–∞—Ä–∏–π (Diplodiscus) –∏–ª–∏ –¥–∞–∂–µ —Ä–∞–∑–≤–∏—Ç—ã–µ –≥–ª–∞–∑–∞ (—É –ø–∞—Ä–∞–∑–∏—Ç–∞ –º–æ—Ä—Å–∫–∏—Ö —Ä—ã–±).
–ü–æ–ª–æ–≤–∞—è —Å–∏—Å—Ç–µ–º–∞ —É —Ç—Ä–µ–º–∞—Ç–æ–¥, –∫–∞–∫ –ø—Ä–∞–≤–∏–ª–æ, –≥–µ—Ä–º–∞—Ñ—Ä–æ–¥–∏—Ç–Ω–∞. –ú—É–∂—Å–∫–∏–µ –ø–æ–ª–æ–≤—ã–µ –æ—Ä–≥–∞–Ω—ã –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω—ã —Å–µ–º–µ–Ω–Ω–∏–∫–∞–º–∏, —Å–µ–º—è–ø—Ä–æ–≤–æ–¥–∞–º–∏, —Å–µ–º–µ–Ω–Ω—ã–º –ø—É–∑—ã—Ä—å–∫–æ–º –∏ —Å–æ–≤–æ–∫—É–ø–∏—Ç–µ–ª—å–Ω—ã–º –æ—Ä–≥–∞–Ω–æ–º.
–°–µ–º–µ–Ω–Ω–∏–∫–∏, —á–∞—â–µ –≤—Å–µ–≥–æ –≤ —á–∏—Å–ª–µ –¥–≤—É—Ö, —Ä–∞—Å–ø–æ–ª–∞–≥–∞—é—Ç—Å—è –æ–¥–∏–Ω –∑–∞ –¥—Ä—É–≥–∏–º –∏–ª–∏ —Ä—è–¥–æ–º, –Ω–∞ –æ–¥–Ω–æ–º —É—Ä–æ–≤–Ω–µ. –ò–Ω–æ–≥–¥–∞ –∏–º–µ–µ—Ç—Å—è –æ–¥–∏–Ω —Å–µ–º–µ–Ω–Ω–∏–∫ (—Å–µ–º. Monorchidae) –∏–ª–∏ –∏—Ö –º–æ–∂–µ—Ç –±—ã—Ç—å –º–Ω–æ–∂–µ—Å—Ç–≤–æ (—Å–µ–º.Orchipedidae, Schistosomatidae –∏ –¥—Ä.). –°–µ–º–µ–Ω–Ω–∏–∫–∏ –º–æ–≥—É—Ç –±—ã—Ç—å —Ü–µ–ª—å–Ω–æ–∫—Ä–æ–π–Ω—ã–º–∏ –∏–ª–∏ –ª–æ–ø–∞—Å—Ç–Ω—ã–º–∏, –∞ —É –∫—Ä—É–ø–Ω—ã—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ (—Å–µ–º. Fasciolidae) –Ω–µ—Ä–µ–¥–∫–æ —Ä–∞—Å—Å–µ—á–µ–Ω—ã –Ω–∞ –º–Ω–æ–∂–µ—Å—Ç–≤–æ –¥–æ–ª–µ–π –∏ –¥–∞–∂–µ –¥—Ä–µ–≤–æ–≤–∏–¥–Ω—ã. –û—Ç —Å–µ–º–µ–Ω–Ω–∏–∫–æ–≤ –æ—Ç—Ö–æ–¥—è—Ç —Å–µ–º—è–ø—Ä–æ–≤–æ–¥—ã, –∫–æ—Ç–æ—Ä—ã–µ, —Å–ª–∏–≤–∞—è—Å—å, –æ–±—Ä–∞–∑—É—é—Ç —Å–µ–º–µ–Ω–Ω–æ–π –ø—É–∑—ã—Ä–µ–∫, –ø–æ—Å–ª–µ–¥–Ω–∏–π —Å–≤–æ–±–æ–¥–Ω–æ –ª–µ–∂–∏—Ç –≤ –ø–∞—Ä–µ–Ω—Ö–∏–º–µ –∏–ª–∏ –∑–∞–∫–ª—é—á–µ–Ω –≤–Ω—É—Ç—Ä–∏ —Å—É–º–∫–∏ —Ü–∏—Ä—Ä—É—Å–∞. –í –ø–æ–ª–æ–≤–æ–π –±—É—Ä—Å–µ (–≥—Ä—É—à–µ–≤–∏–¥–Ω—ã–π –º–µ—à–æ–∫ —Å —Å–∏–ª—å–Ω—ã–º–∏ –º—É—Å–∫—É–ª—å–Ω—ã–º–∏ —Å—Ç–µ–Ω–∫–∞–º–∏) –Ω–∞—Ö–æ–¥–∏—Ç—Å—è —Å–æ–≤–æ–∫—É–ø–∏—Ç–µ–ª—å–Ω—ã–π –æ—Ä–≥–∞–Ω - —Ü–∏—Ä—Ä—É—Å –∏–ª–∏ –ø–µ–Ω–∏—Å, –ø—Ä–æ–Ω–∏–∑–∞–Ω–Ω—ã–π —Å–µ–º—è–∏–∑–≤–µ—Ä–≥–∞—Ç–µ–ª—å–Ω—ã–º –∫–∞–Ω–∞–ª–æ–º.
В состав женской половой системы входят яичник, желточные железы и их протоки, тельце Мелиса, оотип, Лауреров канал, матка, семяприемник. Яичник обычно один, чаще шаровидной формы, реже лопастной или разветвленный, располагается впереди семенников или между ними. Короткий яйцевод ведет в оотип – небольшую камеру, где осуществляются процессы оплодотворения и формирования сложного яйца. Сюда же открываются особые железы (тельце Мелиса), которые выделяют жидкость, омывающую оотип и матку и облегчающую скольжение яиц по половым путям. В оотип впадают канал семяприемника, проток желточного резервуара. Через Лауреров канал оотип сообщается с поверхностью тела. По каналу удаляется избыток желточных клеток, также он может выполнять функцию вагины. От оотипа берет начало сильно извитая труба – матка, в которую по мере формирования поступают сложные яйца. Матка открывается наружу рядом с мужским половым отверстием. Желточники гроздевидные, реже компактные, располагаются по бокам тела трематоды. Их протоки сливаются, образуя желточный резервуар. Желточные клетки поступают в желточный резервуар, из него – в оотип, где и входят в состав сложного яйца. Трематоды из подотряда Schistosomatata - раздельнополые паразиты.
–Ø–π—Ü–æ. –ó—Ä–µ–ª—ã–µ —è–π—Ü–∞ —Ç—Ä–µ–º–∞—Ç–æ–¥, –æ–¥–µ—Ç—ã–µ –ø–ª–æ—Ç–Ω–æ–π –æ–±–æ–ª–æ—á–∫–æ–π, –Ω–∞–∑—ã–≤–∞—é—Ç —Å–ª–æ–∂–Ω—ã–º–∏, —Ç.–∫. –æ–Ω–∏ —Å–ª–∞–≥–∞—é—Ç—Å—è –∏–∑ —Ä–∞–∑–Ω–æ—Ä–æ–¥–Ω—ã—Ö –∫–ª–µ—Ç–æ—á–Ω—ã—Ö —ç–ª–µ–º–µ–Ω—Ç–æ–≤. –§–æ—Ä–º–∞ —Å–ª–æ–∂–Ω–æ–≥–æ —è–π—Ü–∞ –æ–±—ã—á–Ω–æ —É–¥–ª–∏–Ω–µ–Ω–Ω–æ-–æ–≤–∞–ª—å–Ω–∞—è; —É –Ω–µ–∫–æ—Ç–æ—Ä—ã—Ö –≤–∏–¥–æ–≤ —è–π—Ü–∞ –∏–º–µ—é—Ç –Ω–µ–ø—Ä–∞–≤–∏–ª—å–Ω—É—é —Ñ–æ—Ä–º—É, —Å–Ω–∞–±–∂–µ–Ω—ã —à–∏–ø–∞–º–∏ –∏–ª–∏ —Ñ–∏–ª–∞–º–µ–Ω—Ç–∞–º–∏ (–æ—Ç—Ä–æ—Å—Ç–∫–∞–º–∏). –ù–∞ –æ–¥–Ω–æ–º –∏–∑ –ø–æ–ª—é—Å–æ–≤ —è–π—Ü–∞ –∏–º–µ–µ—Ç—Å—è –∫—Ä—ã—à–µ—á–∫–∞. –í –º–æ–º–µ–Ω—Ç –≤—ã–ª—É–ø–ª–µ–Ω–∏—è –∫—Ä—ã—à–µ—á–∫–∞ –æ—Ç–∫–∏–¥—ã–≤–∞–µ—Ç—Å—è, –¥–∞–≤–∞—è –ª–∏—á–∏–Ω–∫–µ –≤–æ–∑–º–æ–∂–Ω–æ—Å—Ç—å –≤—ã—Ö–æ–¥–∞ –Ω–∞—Ä—É–∂—É. –Ø–π—Ü–∞ —Ç—Ä–µ–º–∞—Ç–æ–¥ –º–æ–≥—É—Ç –±—ã—Ç—å –ø—Ä–æ–∑—Ä–∞—á–Ω—ã–º–∏, —Å–≤–µ—Ç–ª–æ-–∫–æ—Ä–∏—á–Ω—ã–º–∏, —Å–≤–µ—Ç–ª–æ-–∂–µ–ª—Ç—ã–º–∏, —Ç–µ–º–Ω–æ-–∫–æ—Ä–∏—á–Ω–µ–≤—ã–º–∏, —á–µ—Ä–Ω—ã–º–∏.
–í–Ω—É—Ç—Ä–∏ —è–π—Ü–∞ —Å–æ–¥–µ—Ä–∂–∞—Ç—Å—è –æ–ø–ª–æ–¥–æ—Ç–≤–æ—Ä–µ–Ω–Ω–∞—è —è–π—Ü–µ–∫–ª–µ—Ç–∫–∞ –∏ –∂–µ–ª—Ç–æ—á–Ω—ã–µ –∫–ª–µ—Ç–∫–∏. –ñ–µ–ª—Ç–æ—á–Ω—ã–µ –∫–ª–µ—Ç–∫–∏ –ø–æ—Å—Ç–∞–≤–ª—è—é—Ç –Ω–µ–æ–±—Ö–æ–¥–∏–º—ã–π –¥–ª—è —Ä–∞–∑–≤–∏—Ç–∏—è –∑–∞—Ä–æ–¥—ã—à–∞ —ç–Ω–µ—Ä–≥–µ—Ç–∏—á–µ—Å–∫–∏–π –º–∞—Ç–µ—Ä–∏–∞–ª (–∂–µ–ª—Ç–æ–∫), —Ç.–∫. —Å–∞–º–∞ —è–π—Ü–µ–∫–ª–µ—Ç–∫–∞ —Å–æ–≤–µ—Ä—à–µ–Ω–Ω–æ –ª–∏—à–µ–Ω–∞ –ø–∏—Ç–∞—Ç–µ–ª—å–Ω—ã—Ö –≤–µ—â–µ—Å—Ç–≤.
Оболочка яйца трематод - защитное образование; она состоит из четырех хорошо различимых слоев. Снаружи яйцо одето очень тонкой бесцветной оболочкой, за которой следует более плотная оболочка темно-коричневого цвета или золотисто-желтого. Третья тонкая оболочка плотно прилегает к четвертой (внутренней). Три наружные оболочки служат для защиты зародыша от механических повреждений, тогда как четвертая – внутренняя - защищает его от химических воздействий и является полупроницаемой.
–ú–∏—Ä–∞—Ü–∏–¥–∏–π. –≠–º–±—Ä–∏–æ–Ω–∞–ª—å–Ω–æ–µ —Ä–∞–∑–≤–∏—Ç–∏–µ —Ç—Ä–µ–º–∞—Ç–æ–¥ —Ö–∞—Ä–∞–∫—Ç–µ—Ä–∏–∑—É–µ—Ç—Å—è –ø–æ–ª–Ω—ã–º –Ω–µ—Ä–∞–≤–Ω–æ–º–µ—Ä–Ω—ã–º –∏ –Ω–µ–ø—Ä–∞–≤–∏–ª—å–Ω—ã–º –¥—Ä–æ–±–ª–µ–Ω–∏–µ–º. –ü–µ—Ä–≤–æ–µ –¥–µ–ª–µ–Ω–∏–µ –æ–ø–ª–æ–¥–æ—Ç–≤–æ—Ä–µ–Ω–Ω–æ–π —è–π—Ü–µ–∫–ª–µ—Ç–∫–∏ –ø—Ä–∏–≤–æ–¥–∏—Ç –∫ –æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏—é –¥–≤—É—Ö –Ω–µ–æ–¥–∏–Ω–∞–∫–æ–≤—ã—Ö –ø–æ –≤–µ–ª–∏—á–∏–Ω–µ –±–ª–∞—Å—Ç–æ–º–µ—Ä–æ–≤: –±–æ–ª–µ–µ –∫—Ä—É–ø–Ω—ã–π - —ç–∫—Ç–æ–¥–µ—Ä–º–∞–ª—å–Ω—ã–π, –º–µ–Ω—å—à–∏–π - –ø—Ä–µ–¥—Å—Ç–∞–≤–ª—è–µ—Ç —Å–æ–±–æ–π –±—É–¥—É—â–∏–π –ø–æ–ª–æ–≤–æ–π –∑–∞—á–∞—Ç–æ–∫ –∏ –Ω–∞–∑—ã–≤–∞–µ—Ç—Å—è –ø—Ä–æ–ø–∞–≥–∞—Ç–∏–≤–Ω—ã–º. –í –¥–∞–ª—å–Ω–µ–π—à–µ–º —Ç–µ–º–ø –¥—Ä–æ–±–ª–µ–Ω–∏—è –¥–≤—É—Ö –ø–µ—Ä–≤—ã—Ö –±–ª–∞—Å—Ç–æ–º–µ—Ä–æ–≤ –Ω–µ–æ–¥–∏–Ω–∞–∫–æ–≤. –≠–∫—Ç–æ–¥–µ—Ä–º–∞–ª—å–Ω—ã–π –±–ª–∞—Å—Ç–æ–º–µ—Ä, –ø—Ä–µ–¥—Å—Ç–∞–≤–ª—è—é—â–∏–π —Å–æ–±–æ–π —Å–æ–º–∞—Ç–∏—á–µ—Å–∫–∏–π –∑–∞—á–∞—Ç–æ–∫, —Å—Ä–∞–∑—É –∂–µ –∏–Ω—Ç–µ–Ω—Å–∏–≤–Ω–æ –¥–µ–ª–∏—Ç—Å—è, –≤ —Ä–µ–∑—É–ª—å—Ç–∞—Ç–µ —á–µ–≥–æ –æ–±—Ä–∞–∑—É—é—Ç—Å—è —Å–æ–º–∞—Ç–∏—á–µ—Å–∫–∏–µ —Å—Ç—Ä—É–∫—Ç—É—Ä—ã –º–∏—Ä–∞—Ü–∏–¥–∏—è, –ª–∏–±–æ —á–∞—Å—Ç—å –∏—Ö.
–í —Å—Ç—Ä–æ–µ–Ω–∏–∏ –º–∏—Ä–∞—Ü–∏–¥–∏–µ–≤ —Ä–∞–∑–Ω—ã—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ –∏–º–µ—é—Ç—Å—è —Å–ø–µ—Ü–∏—Ñ–∏—á–µ—Å–∫–∏–µ —á–µ—Ä—Ç—ã, —Ö–∞—Ä–∞–∫—Ç–µ—Ä–Ω—ã–µ –¥–ª—è –æ—Ç–¥–µ–ª—å–Ω—ã—Ö —Å–∏—Å—Ç–µ–º–∞—Ç–∏—á–µ—Å–∫–∏—Ö –≥—Ä—É–ø–ø. –¢–µ–º –Ω–µ –º–µ–Ω–µ–µ –≤ –º–æ—Ä—Ñ–æ–ª–æ–≥–∏–∏ –º–∏—Ä–∞—Ü–∏–¥–∏–µ–≤ —Å—Ç–æ–ª—å–∫–æ —Å—Ö–æ–¥–Ω–æ–≥–æ, —á—Ç–æ –≤–ø–æ–ª–Ω–µ –º–æ–∂–Ω–æ –¥–∞—Ç—å –æ–±—â—É—é –∏—Ö —Ö–∞—Ä–∞–∫—Ç–µ—Ä–∏—Å—Ç–∏–∫—É. –Ý–∞–∑–º–µ—Ä—ã –º–∏—Ä–∞—Ü–∏–¥–∏–µ–≤ –∫–æ–ª–µ–±–ª—é—Ç—Å—è –æ—Ç 0,02 –¥–æ 0,34 –º–º. –¢–µ–ª–æ —Ü–∏–ª–∏–Ω–¥—Ä–∏—á–µ—Å–∫–æ–π —Ñ–æ—Ä–º—ã, –≤ –ø–µ—Ä–µ–¥–Ω–µ–π —Ç—Ä–µ—Ç–∏ –æ–Ω–æ —Ä–∞—Å—à–∏—Ä–µ–Ω–æ. –ù–∞ —Å–∞–º–æ–º –ø–µ—Ä–µ–¥–Ω–µ–º –∫–æ–Ω—Ü–µ —Ç–µ–ª–∞ –Ω–∞—Ö–æ–¥–∏—Ç—Å—è –ø–æ–¥–≤–∏–∂–Ω—ã–π, —Å–ø–æ—Å–æ–±–Ω—ã–π –≤—ã–≤–æ—Ä–∞—á–∏–≤–∞—Ç—å—Å—è, –º—É—Å–∫—É–ª—å–Ω—ã–π —Ö–æ–±–æ—Ç–æ–∫, –∏–≥—Ä–∞—é—â–∏–π –≤–∞–∂–Ω—É—é —Ä–æ–ª—å –≤ –ø—Ä–æ—Ü–µ—Å—Å–µ –ø—Ä–æ–Ω–∏–∫–Ω–æ–≤–µ–Ω–∏—è –ª–∏—á–∏–Ω–∫–∏ –≤ —Ç–∫–∞–Ω–∏ –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω–æ–≥–æ —Ö–æ–∑—è–∏–Ω–∞.
–ü–æ–∫—Ä–æ–≤—ã –º–∏—Ä–∞—Ü–∏–¥–∏—è –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω—ã –∫—Ä—É–ø–Ω—ã–º–∏ –ø–ª–æ—Å–∫–∏–º–∏ –∫–ª–µ—Ç–∫–∞–º–∏ (—ç–ø–∏—Ç–µ–ª–∏–∞–ª—å–Ω—ã–µ –ø–ª–∞—Å—Ç–∏–Ω–∫–∏), –∫–æ—Ç–æ—Ä—ã–µ –Ω–µ—Å—É—Ç —Ä–µ—Å–Ω–∏—á–∫–∏, —Ä–∞—Å–ø–æ–ª–æ–∂–µ–Ω–Ω—ã–µ –ø—Ä–∞–≤–∏–ª—å–Ω—ã–º–∏ –ø—Ä–æ–¥–æ–ª—å–Ω—ã–º–∏ —Ä—è–¥–∞–º–∏. –ö–∞–∂–¥–∞—è —Ä–µ—Å–Ω–∏—á–∫–∞ —Å–æ–µ–¥–∏–Ω–µ–Ω–∞ —Å –±–∞–∑–∞–ª—å–Ω—ã–º –∑–µ—Ä–Ω–æ–º. –ü–æ–¥ —ç–ø–∏—Ç–µ–ª–∏–∞–ª—å–Ω—ã–º–∏ –ø–ª–∞—Å—Ç–∏–Ω–∫–∞–º–∏ –ª–µ–∂–∞—Ç –∫–ª–µ—Ç–∫–∏ –≥–∏–ø–æ–¥–µ—Ä–º—ã.
–í –ø–µ—Ä–µ–¥–Ω–µ–π —á–∞—Å—Ç–∏ —Ç–µ–ª–∞ –º–∏—Ä–∞—Ü–∏–¥–∏–µ–≤ —Å–æ—Å—Ä–µ–¥–æ—Ç–æ—á–µ–Ω—ã —Ä–∞–∑–ª–∏—á–Ω—ã–µ –ø–æ —Å–≤–æ–µ–º—É —Ö–∞—Ä–∞–∫—Ç–µ—Ä—É –∏ —Ñ—É–Ω–∫—Ü–∏—è–º –∂–µ–ª–µ–∑–∏—Å—Ç—ã–µ –∫–ª–µ—Ç–∫–∏. –ö—Ä—É–ø–Ω–∞—è –∞–ø–∏–∫–∞–ª—å–Ω–∞—è –∂–µ–ª–µ–∑–∞, –ø—Ä–æ—Ç–æ–∫–∏ –∫–æ—Ç–æ—Ä–æ–π –æ—Ç–∫—Ä—ã–≤–∞—é—Ç—Å—è –Ω–∞ –ø–µ—Ä–µ–¥–Ω–µ–º –∫–æ–Ω—Ü–µ —Ç–µ–ª–∞, –ø—Ä–µ–¥—Å—Ç–∞–≤–ª—è–µ—Ç —Å–æ–±–æ–π —Å–∏–Ω—Ü–∏—Ç–∏–π —Å —á–µ—Ç—ã—Ä—å–º—è —è–¥—Ä–∞–º–∏. –≠—Ç–∞ –∂–µ–ª–µ–∑–∞- –æ—Ä–≥–∞–Ω –ø—Ä–æ–Ω–∏–∫–Ω–æ–≤–µ–Ω–∏—è. –ü–æ –æ–±–µ —Å—Ç–æ—Ä–æ–Ω—ã –æ—Ç –∞–ø–∏–∫–∞–ª—å–Ω–æ–π –∂–µ–ª–µ–∑—ã —Ä–∞—Å–ø–æ–ª–∞–≥–∞—é—Ç—Å—è –æ–¥–Ω–æ–∫–ª–µ—Ç–æ—á–Ω—ã–µ –∂–µ–ª–µ–∑—ã –≤ —á–∏—Å–ª–µ –¥–≤—É—Ö-—Ç—Ä–µ—Ö –ø–∞—Ä, –ø—Ä–æ—Ç–æ–∫–∏ –∫–æ—Ç–æ—Ä—ã—Ö –æ—Ç—Ä—ã–≤–∞—é—Ç—Å—è –Ω–∞ –ø–µ—Ä–µ–¥–Ω–µ–º –∫–æ–Ω—Ü–µ —Ç–µ–ª–∞ –º–∏—Ä–∞—Ü–∏–¥–∏—è. –≠—Ç–∏ –∂–µ–ª–µ–∑—ã –∏–≥—Ä–∞—é—Ç –≤–∞–∂–Ω—É—é —Ä–æ–ª—å –≤ –ø—Ä–æ—Ü–µ—Å—Å–µ –≤—ã–ª—É–ø–ª–µ–Ω–∏—è –º–∏—Ä–∞—Ü–∏–¥–∏–µ–≤ –∏–∑ —è–π—Ü–∞.
–ü–∞—Ä—Ç–µ–Ω–æ–≥–µ–Ω–µ—Ç–∏—á–µ—Å–∫–∏–µ –ø–æ–∫–æ–ª–µ–Ω–∏—è —Ç—Ä–µ–º–∞—Ç–æ–¥. –ú–∏—Ä–∞—Ü–∏–¥–∏–π –ø–æ—Å–ª–µ –ø—Ä–æ–Ω–∏–∫–Ω–æ–≤–µ–Ω–∏—è –≤ –æ—Ä–≥–∞–Ω–∏–∑–º —Ö–æ–∑—è–∏–Ω–∞ –ø—Ä–µ—Ç–µ—Ä–ø–µ–≤–∞–µ—Ç –º–µ—Ç–∞–º–æ—Ä—Ñ–æ–∑ –∏ –ø—Ä–µ–≤—Ä–∞—â–∞–µ—Ç—Å—è –≤ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫—É—é —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—É. –≠—Ç–æ—Ç –º–µ—Ç–∞–º–æ—Ä—Ñ–æ–∑ —Ä–µ–≥—Ä–µ—Å—Å–∏–≤–µ–Ω, —Ç.–∫. —Å–≤—è–∑–∞–Ω —Å —É—Ç—Ä–∞—Ç–æ–π —Ä—è–¥–∞ –ª–∏—á–∏–Ω–æ—á–Ω—ã—Ö –ø—Ä–∏–∑–Ω–∞–∫–æ–≤ –∏ –ø—Ä–∏–≤–æ–¥–∏—Ç –∫ –∑–Ω–∞—á–∏—Ç–µ–ª—å–Ω–æ–º—É —É–ø—Ä–æ—â–µ–Ω–∏—é –æ—Ä–≥–∞–Ω–∏–∑–∞—Ü–∏–∏ –ø–∞—Ä–∞–∑–∏—Ç–∞. –ï—â–µ –≤ –ø—Ä–æ—Ü–µ—Å—Å–µ –≤–Ω–µ–¥—Ä–µ–Ω–∏—è (Fasciola hepatica) –∏–ª–∏ –≤ –ø–µ—Ä–≤—ã–µ —á–∞—Å—ã –ø–æ—Å–ª–µ –ø–æ–ø–∞–¥–∞–Ω–∏—è –≤ –æ—Ä–≥–∞–Ω–∏–∑–º –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω–æ–≥–æ —Ö–æ–∑—è–∏–Ω–∞ (Paramphistomum microbothrium, Schistosoma mansoni) –º–∏—Ä–∞—Ü–∏–¥–∏–π —Å–±—Ä–∞—Å—ã–≤–∞–µ—Ç —ç–ø–∏—Ç–µ–ª–∏–∞–ª—å–Ω—ã–µ –ø–ª–∞—Å—Ç–∏–Ω–∫–∏, –æ–±–Ω–∞–∂–∞–µ—Ç—Å—è —Å—É–±—ç–ø–∏—Ç–µ–ª–∏–∞–ª—å–Ω—ã–π —Å–ª–æ–π, –ø–æ—Å–ª–µ–¥–Ω–∏–π —Å—Ç–∞–Ω–æ–≤–∏—Ç—Å—è —Å—Ç–µ–Ω–∫–æ–π –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã. –õ–∏—á–∏–Ω–æ—á–Ω–∞—è –∞–ø–∏–∫–∞–ª—å–Ω–∞—è –∂–µ–ª–µ–∑–∞, —Å–µ–∫—Ä–µ—Ç –∫–æ—Ç–æ—Ä–æ–π –æ–±–µ—Å–ø–µ—á–∏–≤–∞–µ—Ç –≤–æ–∑–º–æ–∂–Ω–æ—Å—Ç—å –ø–µ—Ä–µ–¥–≤–∏–∂–µ–Ω–∏—è –ø–∞—Ä–∞–∑–∏—Ç–∞ –ø–æ —Ç–∫–∞–Ω—è–º —Ö–æ–∑—è–∏–Ω–∞, —Ñ—É–Ω–∫—Ü–∏–æ–Ω–∏—Ä—É–µ—Ç –¥–æ –¥–æ—Å—Ç–∏–∂–µ–Ω–∏—è –ª–∏—á–∏–Ω–∫–æ–π –º–µ—Å—Ç–∞ —Å–≤–æ–µ–≥–æ –æ–±–∏—Ç–∞–Ω–∏—è, –ø–æ—Å–ª–µ —á–µ–≥–æ –æ–Ω–∞ —Ä–∞–∑—Ä—É—à–∞–µ—Ç—Å—è. –ó–∞—Ç–µ–º –ø—Ä–æ–∏—Å—Ö–æ–¥–∏—Ç —Ä–µ–¥—É–∫—Ü–∏—è –æ—Ä–≥–∞–Ω–æ–≤ —á—É–≤—Å—Ç–≤, —É–ø—Ä–æ—â–∞–µ—Ç—Å—è —Å—Ç—Ä–æ–µ–Ω–∏–µ –Ω–µ—Ä–≤–Ω–æ–π —Å–∏—Å—Ç–µ–º—ã, —É –Ω–µ–∫–æ—Ç–æ—Ä—ã—Ö –≤–∏–¥–æ–≤ —Ä–∞–∑—Ä—É—à–∞–µ—Ç—Å—è –º–æ–∑–≥–æ–≤–æ–π –≥–∞–Ω–≥–ª–∏–π, –∏—Å—á–µ–∑–∞—é—Ç –≥–ª–∞–∑–∞. –ò–∑ –æ—Ä–≥–∞–Ω–æ–≤ –º–∏—Ä–∞—Ü–∏–¥–∏—è –≤ –∑—Ä–µ–ª–æ–π –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–µ —Å–æ—Ö—Ä–∞–Ω—è—é—Ç—Å—è —è–∏—á–Ω–∏–∫ (–∑–∞—Ä–æ–¥—ã—à–µ–≤—ã–µ –∫–ª–µ—Ç–∫–∏) –∏ –ø—Ä–æ—Ç–æ–Ω–µ—Ñ—Ä–∏–¥–∏–∏. –ú–µ—Ç–∞–º–æ—Ä—Ñ–æ–∑ –º–∏—Ä–∞—Ü–∏–¥–∏—è –∑–∞–≤–µ—Ä—à–∞–µ—Ç—Å—è –≤ —Ç–µ—á–µ–Ω–∏–µ –¥–≤—É—Ö-–ø—è—Ç–∏ –¥–Ω–µ–π, –∏–Ω–æ–≥–¥–∞ –Ω–µ—Å–∫–æ–ª—å–∫–æ —á–∞—Å–æ–≤ (Fasciola hepatica).
–ü–æ–ª–Ω–æ—Å—Ç—å—é —Å—Ñ–æ—Ä–º–∏—Ä–æ–≤–∞–Ω–Ω–∞—è –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–∞—è —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–∞ –ø—Ä–µ–¥—Å—Ç–∞–≤–ª—è–µ—Ç —Å–æ–±–æ–π —Ç–æ–Ω–∫–æ—Å—Ç–µ–Ω–Ω—ã–π –æ–∫—Ä—É–≥–ª—ã–π, –≤–µ—Ä–µ—Ç–µ–Ω–æ–æ–±—Ä–∞–∑–Ω—ã–π –∏–ª–∏ –Ω–∏—Ç–µ–≤–∏–¥–Ω—ã–π –º–µ—à–æ–∫ —Å –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã–º–∏ –∫–ª–µ—Ç–∫–∞–º–∏ –∏ —à–∞—Ä–∞–º–∏. –Ý–∞–∑–º–µ—Ä—ã –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–∏—Ö —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç –æ–±—ã—á–Ω–æ –∫–æ–ª–µ–±–ª—é—Ç—Å—è –≤ –ø—Ä–µ–¥–µ–ª–∞—Ö 1-3 –º–º, –Ω–æ —É –Ω–µ–∫–æ—Ç–æ—Ä—ã—Ö –≤–∏–¥–æ–≤ –æ–Ω–∏ –¥–æ—Å—Ç–∏–≥–∞—é—Ç –æ—á–µ–Ω—å –∑–Ω–∞—á–∏—Ç–µ–ª—å–Ω–æ–π –≤–µ–ª–∏—á–∏–Ω—ã (—Å–µ–º. Schistosomatidae –¥–æ 10 –º–º.). –ü–æ –º–µ—Ä–µ —Ä–æ—Å—Ç–∞ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã —Ñ–æ—Ä–º–∞ –µ–µ —Ç–µ–ª–∞ –º–µ–Ω—è–µ—Ç—Å—è.
–ú–∞—Ç–µ—Ä–∏–Ω—Å–∫–∏–µ —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã —á–∞—â–µ –≤—Å–µ–≥–æ –ª–æ–∫–∞–ª–∏–∑—É—é—Ç—Å—è –≤ –ø–æ–ª–æ—Å—Ç–∏ —Ç–µ–ª–∞, —Ö–æ—Ç—è –æ–Ω–∏ –º–æ–≥—É—Ç –≤—Å—Ç—Ä–µ—á–∞—Ç—å—Å—è –≤ —Ä–∞–∑–ª–∏—á–Ω—ã—Ö –æ—Ä–≥–∞–Ω–∞—Ö –º–æ–ª–ª—é—Å–∫–∞, –≤ —á–∞—Å—Ç–Ω–æ—Å—Ç–∏, –≤ –º–∞–Ω—Ç–∏–∏, –ø–µ—á–µ–Ω–∏, —Å—Ç–µ–Ω–∫–µ –∫–∏—à–µ—á–Ω–∏–∫–∞ (–º–µ–∂–¥—É –æ—Å–Ω–æ–≤–∞–Ω–∏–µ–º –∫–∏—à–µ—á–Ω–æ–≥–æ —ç–ø–∏—Ç–µ–ª–∏—è –∏ –ø—Ä–∏–ª–µ–≥–∞—é—â–∏–º–∏ —Ç–∫–∞–Ω—è–º–∏). –ü–æ –º–µ—Ä–µ —Ä–æ—Å—Ç–∞ —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–∞ –æ–±—Ä–∞–∑—É–µ—Ç –≤—ã–ø—è—á–∏–≤–∞–Ω–∏–µ –Ω–∞ –Ω–∞—Ä—É–∂–Ω–æ–π –ø–æ–≤–µ—Ä—Ö–Ω–æ—Å—Ç–∏ –∫–∏—à–µ—á–Ω–∏–∫–∞, –∏–Ω–æ–≥–¥–∞, –ø—Ä–æ—Ä—ã–≤–∞—è –æ–∫—Ä—É–∂–∞—é—â–∏–µ —Ç–∫–∞–Ω–∏, –æ–∫–∞–∑—ã–≤–∞–µ—Ç—Å—è –Ω–µ–ø–æ—Å—Ä–µ–¥—Å—Ç–≤–µ–Ω–Ω–æ –≤ –≥–µ–º–æ—Ü–µ–ª–µ.
–í –æ–≥—Ä–æ–º–Ω–æ–º –±–æ–ª—å—à–∏–Ω—Å—Ç–≤–µ —Å–ª—É—á–∞–µ–≤ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–∏–µ —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã —Ä–∞–∑–º–Ω–æ–∂–∞—é—Ç—Å—è –ø–∞—Ä—Ç–µ–Ω–æ–≥–µ–Ω–µ—Ç–∏—á–µ—Å–∫–∏–º –ø—É—Ç–µ–º. –í –ø–æ–ª–æ—Å—Ç–∏ —Ç–µ–ª–∞ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–∏—Ö —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç –∑–∞–∫–ª—é—á–µ–Ω—ã –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã–µ (–≥–µ–Ω–µ—Ä–∞—Ç–∏–≤–Ω—ã–µ) –∫–ª–µ—Ç–∫–∏, –∫–æ—Ç–æ—Ä—ã–µ –¥–∞—é—Ç –Ω–∞—á–∞–ª–æ —Å–ª–µ–¥—É—é—â–µ–º—É –ø–æ–∫–æ–ª–µ–Ω–∏—é. –ó–∞—Ä–æ–¥—ã—à–µ–≤—ã–µ –∫–ª–µ—Ç–∫–∏ —á–∞—Å—Ç–æ –±—ã–≤–∞—é—Ç –ø—Ä–∏–∫—Ä–µ–ø–ª–µ–Ω—ã –∫ —Å—Ç–µ–Ω–∫–∞–º —Ç–µ–ª–∞ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã —Å –ø–æ–º–æ—â—å—é —Ü–∏—Ç–æ–ø–ª–∞–∑–º–∞—Ç–∏—á–µ—Å–∫–∏—Ö –≤—ã—Ä–æ—Å—Ç–æ–≤, —Å–æ–≤–æ–∫—É–ø–Ω–æ—Å—Ç—å –∫–æ—Ç–æ—Ä—ã—Ö –æ–±—Ä–∞–∑—É–µ—Ç –Ω–µ–∂–Ω—É—é —Å–µ—Ç—å, –ø–æ—Å–ª–µ–¥–Ω—è—è —Å–ª—É–∂–∏—Ç –∫–∞–∫ –±—ã –æ—Å–Ω–æ–≤–æ–π, –Ω–∞ –∫–æ—Ç–æ—Ä–æ–π –ø–æ–∫–æ—è—Ç—Å—è –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã–µ –∫–ª–µ—Ç–∫–∏. –ü–æ –º–µ—Ä–µ —Ä–∞–∑–≤–∏—Ç–∏—è –æ—Å–æ–±–µ–π –¥–æ—á–µ—Ä–Ω–µ–≥–æ –ø–æ–∫–æ–ª–µ–Ω–∏—è —ç—Ç–∞ —Å–µ—Ç—å —Ä–∞–∑—Ä—É—à–∞–µ—Ç—Å—è –∏ –≤ —Å—Ç–∞—Ä—ã—Ö —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–∞—Ö –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã–µ —à–∞—Ä—ã (—ç–º–±—Ä–∏–æ–Ω—ã) —Å–≤–æ–±–æ–¥–Ω–æ –ø–ª–∞–≤–∞—é—Ç –≤ –ø–æ–ª–æ—Å—Ç–∏ —Ç–µ–ª–∞ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã. –Ý–æ—Å—Ç –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã –æ–±—ã—á–Ω–æ —Å–æ–ø—Ä–æ–≤–æ–∂–¥–∞–µ—Ç—Å—è —É–º–Ω–æ–∂–µ–Ω–∏–µ–º —á–∏—Å–ª–∞ –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫. –¢–∞–∫, –µ—Å–ª–∏ –≤ –∑–∞—Ä–æ–¥—ã—à–µ–≤–æ–π –ø–æ–ª–æ—Å—Ç–∏ –º–∏—Ä–∞—Ü–∏–¥–∏—è Trichobilharzia phosellae —Å–æ–¥–µ—Ä–∂–∏—Ç—Å—è –æ–∫–æ–ª–æ 30 –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫, —Ç–æ —É–∂–µ –Ω–∞ —Ç—Ä–µ—Ç–∏–π –¥–µ–Ω—å —Ä–∞–∑–≤–∏—Ç–∏—è –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã —á–∏—Å–ª–æ –∏—Ö –¥–æ—Å—Ç–∏–≥–∞–µ—Ç 170-180; –≤ –ø–æ–ª–æ—Å—Ç–∏ —Ç–µ–ª–∞ –º–∏—Ä–∞—Ü–∏–¥–∏—è Schistosoma mansoni –∑–∞–∫–ª—é—á–µ–Ω–æ 50-100 –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫, —Ç–æ–≥–¥–∞ –∫–∞–∫ –≤ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–µ —á–∏—Å–ª–æ –∏—Ö 200-400. –í –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–∏—Ö —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–∞—Ö –Ω–∏–∑—à–∏—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ (—Å–µ–º. Fasciolidae, Paramphistotidae) —É–º–Ω–æ–∂–µ–Ω–∏—è —á–∏—Å–ª–∞ –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫ –Ω–µ –ø—Ä–∏—Ö–æ–¥–∏—Ç.
–î–æ—á–µ—Ä–Ω–∏–µ –ø–æ–∫–æ–ª–µ–Ω–∏—è –º–æ–≥—É—Ç –±—ã—Ç—å –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω—ã —Ä–µ–¥–∏—è–º–∏ (Paramphistomotata, Fasciolata, Notocotylata, Echinostomatata) –∏–ª–∏ –¥–æ—á–µ—Ä–Ω–∏–º–∏ —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–∞–º–∏ (Strigeata, Plagiorchidae). –ü–æ –º–µ—Ä–µ —Ñ–æ—Ä–º–∏—Ä–æ–≤–∞–Ω–∏—è –æ—Å–æ–±–∏ –¥–æ—á–µ—Ä–Ω–∏–µ –ø–æ–∫–æ–ª–µ–Ω–∏—è –ø–æ–∫–∏–¥–∞—é—Ç –º–∞—Ç–µ—Ä–∏–Ω—Å–∫—É—é —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—É.
–Ý–µ–¥–∏–∏ –æ–±—ã—á–Ω–æ –æ—Å–≤–æ–±–æ–∂–¥–∞—é—Ç—Å—è, —Ä–∞–∑—Ä—ã–≤–∞—è —Å—Ç–µ–Ω–∫—É –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–∏—Ü–∏—Å—Ç—ã, –∫–æ—Ç–æ—Ä–∞—è –ø—Ä–∏ —ç—Ç–æ–º –ø–æ–≥–∏–±–∞–µ—Ç. –ß–∞—â–µ –≤—Å–µ–≥–æ —ç—Ç–æ –ø—Ä–æ–∏—Å—Ö–æ–¥–∏—Ç –Ω–∞ 12-20 –¥–µ–Ω—å —Ä–∞–∑–≤–∏—Ç–∏—è. –í –æ—Ç–¥–µ–ª—å–Ω—ã—Ö —Å–ª—É—á–∞—è—Ö —Å—Ä–æ–∫ –∂–∏–∑–Ω–∏ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã –º–æ–∂–µ—Ç –±—ã—Ç—å —Ä–∞—Å—Ç—è–Ω—É—Ç –¥–æ –Ω–µ—Å–∫–æ–ª—å–∫–∏—Ö –º–µ—Å—è—Ü–µ–≤ (Notocotylus stagnicola) –∏–ª–∏ —Å–æ–∫—Ä–∞—â–µ–Ω –¥–æ –Ω–µ—Å–∫–æ–ª—å–∫–∏—Ö –¥–Ω–µ–π (—Å–µ–º. Echinostomatidae). –ß–∏—Å–ª–æ —Ä–µ–¥–∏–π, —Ñ–æ—Ä–º–∏—Ä—É—é—â–∏—Ö—Å—è –≤ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–µ, –æ–±—ã—á–Ω–æ –Ω–µ –≤–µ–ª–∏–∫–æ: –≤ —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–µ —É Fasciola gigantica –æ—Ç 1–¥–æ 6, —É Notocotylus quinqueserialis ‚Äì 4, —É Paramphistomum phillerouxi -18-20 —Ä–µ–¥–∏–π.
–î–æ—á–µ—Ä–Ω–∏–µ –ø–æ–∫–æ–ª–µ–Ω–∏—è. 1.–Ý–µ–¥–∏–∏, –ø—Ä–µ–¥—Å—Ç–∞–≤–ª—è—é—â–∏–µ —Å–æ–±–æ–π –¥–æ—á–µ—Ä–Ω–µ–µ –ø–æ–∫–æ–ª–µ–Ω–∏–µ, —Ä–∞–∑–≤–∏–≤–∞—é—Ç—Å—è –≤ –ø–æ–ª–æ—Å—Ç–∏ —Ç–µ–ª–∞ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã. –° —Å–∞–º–æ–≥–æ –Ω–∞—á–∞–ª–∞ —Ä–∞–∑–≤–∏—Ç–∏—è —ç–º–±—Ä–∏–æ–Ω–æ–≤ —Ä–µ–¥–∏–π –≤ –Ω–∏—Ö –º–æ–∂–Ω–æ —Ä–∞–∑–ª–∏—á–∏—Ç—å –∫—Ä—É–ø–Ω—É—é –∑–∞—Ä–æ–¥—ã—à–µ–≤—É—é (–≥–µ–Ω–µ—Ä–∞—Ç–∏–≤–Ω—É—é) –∫–ª–µ—Ç–∫—É, –≤ —Ä–µ–∑—É–ª—å—Ç–∞—Ç–µ –¥–µ–ª–µ–Ω–∏—è –∫–æ—Ç–æ—Ä–æ–π –æ–±—Ä–∞–∑—É–µ—Ç—Å—è –º–æ—Ä—É–ª–æ–ø–æ–¥–æ–±–Ω–æ–µ —Å–∫–æ–ø–ª–µ–Ω–∏–µ –∫–ª–µ—Ç–æ–∫ ‚Äì —è–∏—á–Ω–∏–∫, –∞ —Ç–∞–∫–∂–µ –ø–æ—è–≤–ª—è–µ—Ç—Å—è –∑–∞—á–∞—Ç–æ–∫ –∫–∏—à–µ—á–Ω–∏–∫–∞. –í—Å–µ —Ç–µ–ª–æ –º–æ–ª–æ–¥–æ–π —Ä–µ–¥–∏–∏ –∑–∞–ø–æ–ª–Ω–µ–Ω–æ –ø–∞—Ä–µ–Ω—Ö–∏–º–∞—Ç–æ–∑–Ω—ã–º–∏ –∫–ª–µ—Ç–∫–∞–º–∏. –í –¥–∞–ª—å–Ω–µ–π—à–µ–º –º–µ–∂–¥—É –Ω–∏–º–∏ –æ–±—Ä–∞–∑—É—é—Ç—Å—è —â–µ–ª–∏ –∏ –ª–∞–∫—É–Ω—ã, –≤ —Ä–µ–∑—É–ª—å—Ç–∞—Ç–µ —á–µ–≥–æ —è–∏—á–Ω–∏–∫ –æ–∫–∞–∑—ã–≤–∞–µ—Ç—Å—è –≤ –Ω–µ–±–æ–ª—å—à–æ–π –ø–æ–ª–æ—Å—Ç–∏. –ü–æ –º–µ—Ä–µ —Ä–æ—Å—Ç–∞ —Ä–µ–¥–∏–∏ —ç—Ç–∞ –ø–æ–ª–æ—Å—Ç—å —É–≤–µ–ª–∏—á–∏–≤–∞–µ—Ç—Å—è –∏ –≤ –Ω–µ–µ –ø–æ—Å—Ç—É–ø–∞—é—Ç –æ—Ç–¥–µ–ª—è–µ–º—ã–µ –æ—Ç —è–∏—á–Ω–∏–∫–∞ –ø–∞—Ä—Ç–µ–Ω–æ–≥–µ–Ω–µ—Ç–∏—á–µ—Å–∫–∏–µ —è–π—Ü–∞ (–∑–∞—Ä–æ–¥—ã—à–µ–≤—ã–µ –∫–ª–µ—Ç–∫–∏). –í –ø–æ–ª–æ—Å—Ç–∏ –ø—Ä–æ–∏—Å—Ö–æ–¥–∏—Ç —Ä–∞–∑–≤–∏—Ç–∏–µ –∑–∞—Ä–æ–¥—ã—à–µ–π –¥–æ —Å–ª–µ–¥—É—é—â–µ–≥–æ –ø–æ–∫–æ–ª–µ–Ω–∏—è. –í–∑—Ä–æ—Å–ª—ã–µ —Ä–µ–¥–∏–∏ –∏–º–µ—é—Ç —Ö–æ—Ä–æ—à–æ —Ä–∞–∑–≤–∏—Ç—É—é –ø–∏—â–µ–≤–∞—Ä–∏—Ç–µ–ª—å–Ω—É—é —Å–∏—Å—Ç–µ–º—É (–º—É—Å–∫—É–ª–∏—Å—Ç–∞—è –≥–ª–æ—Ç–∫–∞ –∏ –∫–∏—à–µ—á–Ω–∏–∫).
–ù–∞ –ø–µ—Ä–µ–¥–Ω–µ–º –∫–æ–Ω—Ü–µ —Ç–µ–ª–∞ —Ä–µ–¥–∏–π —Ä–∞—Å–ø–æ–ª–∞–≥–∞–µ—Ç—Å—è –æ–∫—Ä—É–≥–ª–∞—è –∏–ª–∏ –≤—ã—Ç—è–Ω—É—Ç–∞—è –º—É—Å–∫—É–ª–∏—Å—Ç–∞—è –≥–ª–æ—Ç–∫–∞. –û–Ω–∞ –≤–µ–¥–µ—Ç –≤ —Å–ª–µ–ø–æ –∑–∞–º–∫–Ω—É—Ç—ã–π –∫–∏—à–µ—á–Ω–∏–∫. –°—Ç–µ–ø–µ–Ω—å —Ä–∞–∑–≤–∏—Ç–∏—è –∫–∏—à–µ—á–Ω–∏–∫–∞ —Ä–µ–¥–∏–π —è–≤–ª—è–µ—Ç—Å—è –≤–∞–∂–Ω—ã–º —Å–∏—Å—Ç–µ–º–∞—Ç–∏—á–µ—Å–∫–∏–º –ø—Ä–∏–∑–Ω–∞–∫–æ–º.
–ü–æ–ª–æ–≤–∞—è —Å–∏—Å—Ç–µ–º–∞ —Ä–µ–¥–∏–π –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω–∞ –Ω–µ–±–æ–ª—å—à–∏–º –∫–æ–º–ø–∞–∫—Ç–Ω—ã–º —è–∏—á–Ω–∏–∫–æ–º, —Ä–∞—Å–ø–æ–ª–æ–∂–µ–Ω–Ω—ã–º –≤ –∑–∞–¥–Ω–µ–π —Ç—Ä–µ—Ç–∏ —Ç–µ–ª–∞. –ù–∞ –ø—Ä–æ—Ç—è–∂–µ–Ω–∏–∏ –≤—Å–µ–π –∂–∏–∑–Ω–∏ —Ä–µ–¥–∏–∏ –æ—Ç –µ–µ —è–∏—á–Ω–∏–∫–∞ –æ—Ç–¥–µ–ª—è—é—Ç—Å—è –≤—Å–µ –Ω–æ–≤—ã–µ –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã–µ –∫–ª–µ—Ç–∫–∏, –∫–æ—Ç–æ—Ä—ã–µ —Ä–∞—Å—Ç—É—Ç –∏ –Ω–∞—á–∏–Ω–∞—é—Ç –¥—Ä–æ–±–∏—Ç—å—Å—è, –¥–∞–≤–∞—è –Ω–∞—á–∞–ª–æ –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã–º —à–∞—Ä–∞–º. –ü—Ä–æ—Ü–µ—Å—Å –∏—Ö —Ñ–æ—Ä–º–∏—Ä–æ–≤–∞–Ω–∏—è –≤–ø–æ–ª–Ω–µ –∏–¥–µ–Ω—Ç–∏—á–µ–Ω —Ñ–æ—Ä–º–∏—Ä–æ–≤–∞–Ω–∏—é –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã—Ö —à–∞—Ä–æ–≤ –≤ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–µ. –ü–æ –º–µ—Ä–µ —Ä–æ—Å—Ç–∞ –∏ —Å—Ç–∞—Ä–µ–Ω–∏—è —Ä–µ–¥–∏–∏ –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã–µ —à–∞—Ä—ã –∏ —Ä–∞–∑–≤–∏–≤–∞—é—â–∏–µ—Å—è –∑–∞—Ä–æ–¥—ã—à–∏ –¥–æ—á–µ—Ä–Ω–∏—Ö –ø–æ–∫–æ–ª–µ–Ω–∏–π –ø–æ—Å—Ç–µ–ø–µ–Ω–Ω–æ –∑–∞–ø–æ–ª–Ω—è—é—Ç –≤—Å—é –ø–æ–ª–æ—Å—Ç—å —Ç–µ–ª–∞. –ü—Ä–æ–¥—É–∫—Ç–∏–≤–Ω–æ—Å—Ç—å —Ä–µ–¥–∏–π –Ω–µ –≤–µ–ª–∏–∫–∞ (–¥–≤–µ-—á–µ—Ç—ã—Ä–µ –¥–æ—á–µ—Ä–Ω–∏–µ —Ä–µ–¥–∏–∏ –∏–ª–∏ 6-10 —Å—Ñ–æ—Ä–º–∏—Ä–æ–≤–∞–Ω–Ω—ã—Ö —Ü–µ—Ä–∫–∞—Ä–∏–π).
В организме моллюска последовательно развиваются несколько поколений редий. Материнские и дочерние редии отличаются друг от друга не только морфологически, но и биологически. Так, материнские редии могут давать начало только редиям, тогда как дочерние – редиям и, кроме того, отрождают и церкарий.
–ú–µ—Å—Ç–æ–º –ø–∞—Ä–∞–∑–∏—Ç–∏—Ä–æ–≤–∞–Ω–∏—è —Ä–µ–¥–∏–π (–º–∞—Ç–µ—Ä–∏–Ω—Å–∫–∏—Ö –∏ –¥–æ—á–µ—Ä–Ω–∏—Ö) –º–æ–∂–µ—Ç –±—ã—Ç—å –≥–µ–º–æ—Ü–µ–ª—å, –ø–µ—á–µ–Ω—å –º–æ–ª–ª—é—Å–∫–∞. –Ý–µ–¥–∏–∏ –ø–æ–¥–≤–∏–∂–Ω—ã, –æ–¥–Ω–∞–∫–æ —Å –≤–æ–∑—Ä–∞—Å—Ç–æ–º –∏—Ö –ø–æ–¥–≤–∏–∂–Ω–æ—Å—Ç—å —Ä–µ–∑–∫–æ —Å–Ω–∏–∂–∞–µ—Ç—Å—è.
2. –î–æ—á–µ—Ä–Ω—è—è —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–∞. –î–æ—á–µ—Ä–Ω–∏–µ —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã –±–æ–ª–µ–µ –º–µ–ª–∫–∏–µ.
Внутри тела дочерних спороцист многочисленные зародышевые клетки. Они размножаются на протяжении всей жизни спороцисты, давая начало зародышевым шарам, из которых развиваются особи последующего поколения – церкарии. На переднем конце тела спороцист имеется родильная пора, расположенная терминально или субтерминально. Через это отверстие выходят наружу сформированные церкарии. Продуктивность дочерних спороцист выше, чем продуктивность редий.
Церкарии. Свободноживущие личинки трематод – церкарии имеют собственно тело и хвостик, который подвижно сочленен с телом и легко может быть отброшен. Церкарии сем. Fasciolidae, Paramphistomatidae, Echinostomatidae и др. крупные, длина их тела соответствует длине хвоста. Значительное более мелкими размерами характеризуются церкарии высших трематод (Plagiorchidae, Strigeidae).
Тело церкарий имеет листовидную, плоскую или удлиненную (цилиндрическую) форму, несет одну – две присоски; одето тонким тегументом толщиной 1-4 мкм, вооруженный шипиками. Пищеварительная система является уже полностью сформированной и устроена по характерному для класса трематод типу. Она начинается ротовой присоской на переднем конце тела, затем следует короткая предглотка, пищевод и две ветки кишечника. Однако личинки не питаются и никаких следов пищи в их кишечнике обнаружить не удается.
–í—ã–¥–µ–ª–∏—Ç–µ–ª—å–Ω–∞—è —Å–∏—Å—Ç–µ–º–∞ —Ü–µ—Ä–∫–∞—Ä–∏–π –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω–∞ –±–æ–ª—å—à–∏–º —á–∏—Å–ª–æ–º –º–µ—Ä—Ü–∞—Ç–µ–ª—å–Ω—ã—Ö –∫–ª–µ—Ç–æ–∫, —Å–∏—Å—Ç–µ–º–æ–π –∫–∞–Ω–∞–ª—å—Ü–µ–≤, —ç–∫—Å–∫—Ä–µ—Ç–æ—Ä–Ω—ã–º –ø—É–∑—ã—Ä–µ–º, –∫–æ—Ç–æ—Ä—ã–π –æ—Ç–∫—Ä—ã–≤–∞–µ—Ç—Å—è –Ω–∞—Ä—É–∂—É –Ω–µ–ø–∞—Ä–Ω—ã–º –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ–º.
–í —Ç–µ–ª–µ —Ü–µ—Ä–∫–∞—Ä–∏–π –∏–º–µ–µ—Ç—Å—è –±–æ–ª—å—à–æ–µ –∫–æ–ª–∏—á–µ—Å—Ç–≤–æ –∂–µ–ª–µ–∑–∏—Å—Ç—ã—Ö –∫–ª–µ—Ç–æ–∫: —Ü–∏—Å—Ç–æ–≥–µ–Ω–Ω—ã–µ, —Å–ª–∏–∑–∏—Å—Ç—ã–µ –∏ –∂–µ–ª–µ–∑—ã –ø—Ä–æ–Ω–∏–∫–Ω–æ–≤–µ–Ω–∏—è.
–¶–∏—Å—Ç–æ–≥–µ–Ω–Ω—ã–µ –∂–µ–ª–µ–∑—ã - –æ–¥–Ω–æ–∫–ª–µ—Ç–æ—á–Ω—ã–µ –æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏—è, —Ä–∞—Å–ø–æ–ª–æ–∂–µ–Ω–Ω—ã–µ –¥–æ—Ä—Å–∞–ª—å–Ω–æ –≤ –ø–æ–≤–µ—Ä—Ö–Ω–æ—Å—Ç–Ω—ã—Ö —Å–ª–æ—è—Ö –ø–∞—Ä–µ–Ω—Ö–∏–º—ã. –£ —Ü–µ—Ä–∫–∞—Ä–∏–π, –æ—Ç–Ω–æ—Å—è—â–∏—Ö—Å—è –∫ –ø—Ä–∏–º–∏—Ç–∏–≤–Ω—ã–º —Å–µ–º–µ–π—Å—Ç–≤–∞–º —Ç—Ä–µ–º–∞—Ç–æ–¥, —Ü–∏—Å—Ç–æ–≥–µ–Ω–Ω—ã–µ –∂–µ–ª–µ–∑—ã –º–Ω–æ–≥–æ—á–∏—Å–ª–µ–Ω–Ω—ã –∏ —Ä–∞—Å–ø–æ–ª–∞–≥–∞—é—Ç—Å—è —Å–ø–ª–æ—à–Ω—ã–º —Å–ª–æ–µ–º (Notocotylidae, Paramphistomatidae). –£ –Ω–∏–∑—à–∏—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ —Ü–∏—Å—Ç–æ–≥–µ–Ω–Ω—ã–µ –∂–µ–ª–µ–∑—ã —Ü–µ—Ä–∫–∞—Ä–∏–π –Ω–∞–ø–æ–º–∏–Ω–∞—é—Ç —Ä–∞–±–¥–∏—Ç–Ω—ã–µ –∂–µ–ª–µ–∑—ã —Ç—É—Ä–±–µ–ª–ª—è—Ä–∏–∏, –∫–∞–∫ –ø–æ —Å–≤–æ–µ–º—É —Å—Ç—Ä–æ–µ–Ω–∏—é, —Ç–∞–∫ –∏ –ø–æ —Ö–∞—Ä–∞–∫—Ç–µ—Ä—É —Å–µ–∫—Ä–µ—Ç–∞, –∏–º–µ—é—â–µ–≥–æ –≤–∏–¥ –∏–≥–æ–ª–æ—á–µ–∫ –∏ –ø–∞–ª–æ—á–µ–∫. –ö –≤—ã—Å—à–∏—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ —Å–µ–∫—Ä–µ—Ç –æ–±—ã—á–Ω–æ –∑–µ—Ä–Ω–∏—Å—Ç—ã–π, –∫–æ–ª–∏—á–µ—Å—Ç–≤–æ —Ü–∏—Å—Ç–æ–≥–µ–Ω–Ω—ã—Ö –∂–µ–ª–µ–∑ –Ω–µ–≤–µ–ª–∏–∫–æ. –£–º–µ–Ω—å—à–µ–Ω–∏–µ —á–∏—Å–ª–∞ —Ü–∏—Å—Ç–æ–≥–µ–Ω–Ω—ã—Ö –∂–µ–ª–µ–∑ —Å–≤—è–∑–∞–Ω–æ —Å –∏—Å—á–µ–∑–Ω–æ–≤–µ–Ω–∏–µ–º —É –º–Ω–æ–≥–∏—Ö –≤—ã—Å—à–∏—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ (Strigeata, –Ω–µ–∫–æ—Ç–æ—Ä—ã–µ Plagiorchidae) —Å–ø–æ—Å–æ–±–Ω–æ—Å—Ç–∏ –∫ –∏–Ω—Ü–∏—Å—Ç–∏—Ä–æ–≤–∞–Ω–∏—é.
–°–ª–∏–∑–∏—Å—Ç—ã–µ –∂–µ–ª–µ–∑—ã –ø—Ä–µ–¥—Å—Ç–∞–≤–ª—è—é—Ç —Å–æ–±–æ–π –æ–¥–Ω–æ–∫–ª–µ—Ç–æ—á–Ω—ã–µ –æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏—è –Ω–µ–ø—Ä–∞–≤–∏–ª—å–Ω–æ–π –∑–≤–µ–∑–¥—á–∞—Ç–æ–π —Ñ–æ—Ä–º—ã, —Å –∫–æ—Ä–æ—Ç–∫–∏–º–∏ –≤—ã–≤–æ–¥–Ω—ã–º–∏ –ø—Ä–æ—Ç–æ–∫–∞–º–∏; –æ–Ω–∏ —Ä–∞—Å–ø–æ–ª–∞–≥–∞—é—Ç—Å—è –≤–µ–Ω—Ç—Ä–∞–ª—å–Ω–æ, —Ä–µ–∂–µ –¥–æ—Ä—Å–∞–ª—å–Ω–æ –ø–∞—Ä–∞–º–∏ –≤–¥–æ–ª—å —Å—Ä–µ–¥–Ω–µ–π –ª–∏–Ω–∏–∏ —Ç–µ–ª–∞. –ü–µ—Ä–∏–æ–¥ —Å–µ–∫—Ä–µ—Ü–∏–∏ —Å–ª–∏–∑–∏—Å—Ç—ã—Ö –∂–µ–ª–µ–∑ –ø—Ä–∏—Ö–æ–¥–∏—Ç—Å—è –Ω–∞ –≤—Ä–µ–º—è –ø—Ä–µ–±—ã–≤–∞–Ω–∏—è —Ü–µ—Ä–∫–∞—Ä–∏–µ–≤ –≤ –æ—Ä–≥–∞–Ω–∏–∑–º–µ –º–æ–ª–ª—é—Å–∫–∞. –í —Å–æ—Å—Ç–∞–≤ –∏—Ö —Å–µ–∫—Ä–µ—Ç–∞ –≤—Ö–æ–¥—è—Ç –º—É–∫–æ–ø–æ–ª–∏—Å–∞—Ö–∞—Ä–∏–¥—ã, –º—É—Ü–∏–Ω, –≥–µ–∫—Å–∞–∑–∞–º–∏–Ω—ã –∏ –ø—Ä–æ—Ç–µ–∏–Ω—ã. –ú—É–∫–æ–ø–æ–ª–∏—Å–∞—Ö–∞—Ä–∏–¥—ã –∏ –º—É–∫–æ–∏–¥—ã —Å–ª–∏–∑–∏—Å—Ç—ã—Ö –∂–µ–ª–µ–∑ —Å–ª—É–∂–∞—Ç –∏–Ω–≥–∏–±–∏—Ç–æ—Ä–∞–º–∏ –ø—Ä–æ—Ç–µ–æ–ª–∏—Ç–∏—á–µ—Å–∫–∏—Ö —Ñ–µ—Ä–º–µ–Ω—Ç–æ–≤ –º–æ–ª–ª—é—Å–∫–∞. –í–æ –≤—Ä–µ–º—è —Å–≤–æ–±–æ–¥–Ω–æ–π –∂–∏–∑–Ω–∏ —Ü–µ—Ä–∫–∞—Ä–∏–µ–≤, —Å–æ–¥–µ—Ä–∂–∞—â–∞—è—Å—è –≤ —Ä–µ–∑–µ—Ä–≤—É–∞—Ä–∞—Ö, —Å–ª–∏–∑—å —Å–ª–∏–∑–∏—Å—Ç—ã—Ö –∂–µ–ª–µ–∑ –∏—Å–ø–æ–ª—å–∑—É–µ—Ç—Å—è –ª–∏—á–∏–Ω–∫–∞–º–∏ –¥–ª—è –ø—Ä–∏–∫—Ä–µ–ø–ª–µ–Ω–∏—è –∫ —Å—É–±—Å—Ç—Ä–∞—Ç—É –∏ —Ç–µ–ª—É —Ö–æ–∑—è–∏–Ω–∞.
–í–∞–∂–Ω–æ–µ –∑–Ω–∞—á–µ–Ω–∏–µ –≤ –ø—Ä–æ—Ü–µ—Å—Å–µ –∂–∏–∑–Ω–µ–¥–µ—è—Ç–µ–ª—å–Ω–æ—Å—Ç–∏ —Ü–µ—Ä–∫–∞—Ä–∏–µ–≤ –∏–º–µ—é—Ç –∂–µ–ª–µ–∑—ã –ø—Ä–æ–Ω–∏–∫–Ω–æ–≤–µ–Ω–∏—è, –∫–æ—Ç–æ—Ä—ã–µ –∏–Ω–æ–≥–¥–∞ –Ω–∞–∑—ã–≤–∞—é—Ç —Å—Ç–∏–ª–µ—Ç–Ω—ã–º–∏, –∏–ª–∏ –≥–æ–ª–æ–≤–Ω—ã–º–∏ –∂–µ–ª–µ–∑–∞–º–∏. –¶–µ—Ä–∫–∞—Ä–∏–∏ –∏–Ω—Ü–∏—Å—Ç–∏—Ä—É—é—Ç—Å—è –≤–æ –≤–Ω–µ—à–Ω–µ–π —Å—Ä–µ–¥–µ (Notocotylidae, Fasciolidae –∏ –¥—Ä.) –∏–ª–∏, –ø—Ä–æ–Ω–∏–∫–∞—é—Ç –≤ –æ—Ä–≥–∞–Ω–∏–∑–º –¥–æ–ø–æ–ª–Ω–∏—Ç–µ–ª—å–Ω–æ–≥–æ —Ö–æ–∑—è–∏–Ω–∞, –Ω–µ –Ω–∞—Ä—É—à–∞—è —Ü–µ–ª–æ—Å—Ç–Ω–æ—Å—Ç–∏ –µ–≥–æ –ø–æ–∫—Ä–æ–≤–æ–≤ (—Å–µ–º. Echinostomatidae), —Ç.–∫. –ª–∏—à–µ–Ω—ã –∂–µ–ª–µ–∑ —ç—Ç–æ–≥–æ —Ç–∏–ø–∞. –ñ–µ–ª–µ–∑—ã –ø—Ä–æ–Ω–∏–∫–Ω–æ–≤–µ–Ω–∏—è –æ–±—ã—á–Ω–æ –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω—ã –∫—Ä—É–ø–Ω—ã–º–∏ –∫–ª–µ—Ç–∫–∞–º–∏ —Å –±–æ–ª—å—à–∏–º –ø—É–∑—ã—Ä—å–∫–æ–≤–∏–¥–Ω—ã–º —è–¥—Ä–æ–º, –∏—Ö —á–∏—Å–ª–æ –∏ —Ä–∞—Å–ø–æ–ª–æ–∂–µ–Ω–∏–µ –≤ —Ä–∞–∑–Ω—ã—Ö —Å–∏—Å—Ç–µ–º–∞—Ç–∏—á–µ—Å–∫–∏—Ö –≥—Ä—É–ø–ø–∞—Ö —Ü–µ—Ä–∫–∞—Ä–∏–π –¥–æ–≤–æ–ª—å–Ω–æ –ø–æ—Å—Ç–æ—è–Ω–Ω–æ –≤ –ø—Ä–µ–¥–µ–ª–∞—Ö –≤–∏–¥–∞ –∏ –º–æ–∂–µ—Ç —Å–ª—É–∂–∏—Ç—å –¥–∏–∞–≥–Ω–æ—Å—Ç–∏—á–µ—Å–∫–∏–º –ø—Ä–∏–∑–Ω–∞–∫–æ–º. –ß–∞—â–µ –≤—Å–µ–≥–æ –∂–µ–ª–µ–∑—ã –ø—Ä–æ–Ω–∏–∫–Ω–æ–≤–µ–Ω–∏—è —Å–æ–±—Ä–∞–Ω—ã –≤ –¥–≤–µ –≥—Ä—É–ø–ø—ã –∏–∑ –æ–ø—Ä–µ–¥–µ–ª–µ–Ω–Ω–æ–≥–æ —á–∏—Å–ª–∞ –∫–ª–µ—Ç–æ–∫, —Ä–∞—Å–ø–æ–ª–∞–≥–∞—é—â–∏—Ö—Å—è –≤–ø–µ—Ä–µ–¥–∏ –±—Ä—é—à–Ω–æ–π –ø—Ä–∏—Å–æ—Å–∫–∏, –ø–æ–∑–∞–¥–∏ –µ–µ –∏–ª–∏ –ø–æ –æ–±–µ–∏–º —Å—Ç–æ—Ä–æ–Ω–∞–º.
–¶–µ–Ω—Ç—Ä–∞–ª—å–Ω–∞—è –Ω–µ—Ä–≤–Ω–∞—è —Å–∏—Å—Ç–µ–º–∞ —Ü–µ—Ä–∫–∞—Ä–∏–π –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω–∞ –ø–µ—Ä–µ–¥–Ω–∏–º –≥–∞–Ω–≥–ª–∏–µ–º, —Ä–∞—Å–ø–æ–ª–æ–∂–µ–Ω–Ω—ã–º –≤ –ø–µ—Ä–µ–¥–Ω–µ–π —Ç—Ä–µ—Ç–∏ —Ç–µ–ª–∞, –æ–±—ã—á–Ω–æ –Ω–∞ —É—Ä–æ–≤–Ω–µ –≥–ª–æ—Ç–∫–∏. –û—Ç –≥–∞–Ω–≥–ª–∏—è –æ—Ç—Ö–æ–¥—è—Ç –Ω–µ—Å–∫–æ–ª—å–∫–æ –Ω–µ—Ä–≤–Ω—ã—Ö —Ç—è–∂–µ–π –∫ –∑–∞–¥–Ω–µ–º—É –∫–æ–Ω—Ü—É, –∏–∑ –Ω–∏—Ö –Ω–∞–∏–±–æ–ª–µ–µ —Ä–∞–∑–≤–∏—Ç—ã –¥–≤–∞ –≤–µ–Ω—Ç—Ä–∞–ª—å–Ω—ã—Ö —Å—Ç–≤–æ–ª–∞, —Ç–æ–≥–¥–∞ –∫–∞–∫ –ª–∞—Ç–µ—Ä–∞–ª—å–Ω—ã–µ –∏ –¥–æ—Ä—Å–∞–ª—å–Ω—ã–µ –ø–ª–æ—Ö–æ –∑–∞–º–µ—Ç–Ω—ã.
–ò–∑ –æ—Ä–≥–∞–Ω–æ–≤ —á—É–≤—Å—Ç–≤ —É —Ü–µ—Ä–∫–∞—Ä–∏–π –∏–º–µ—é—Ç—Å—è –≥–ª–∞–∑–∞ –∏ —Å–µ–Ω—Å–∏–ª–ª—ã. –ì–ª–∞–∑–∞ —Ç–µ—Å–Ω–æ —Å–≤—è–∑–∞–Ω—ã —Å –º–æ–∑–≥–æ–≤—ã–º –≥–∞–Ω–≥–ª–∏–µ–º; —Å–µ–Ω—Å–∏–ª–ª—ã, —Ä–∞—Å–ø–æ–ª–æ–∂–µ–Ω–Ω—ã–µ –Ω–∞ –ø–æ–≤–µ—Ä—Ö–Ω–æ—Å—Ç–∏ —Ç–µ–ª–∞ —Ü–µ—Ä–∫–∞—Ä–∏–π, –Ω–µ—Å—É—Ç —Ç–æ–Ω–∫–∏–µ –ø–ª–∞–∑–º–∞—Ç–∏—á–µ—Å–∫–∏–µ –≤–æ–ª–æ—Å–∏–∫–∏, —Å–≤—è–∑–∞–Ω–Ω—ã–µ —Å –Ω–µ—Ä–≤–Ω—ã–º–∏ –æ–∫–æ–Ω—á–∞–Ω–∏—è–º–∏. –ù–∞–∏–±–æ–ª—å—à–µ–µ —á–∏—Å–ª–æ —Å–µ–Ω—Å–∏–ª–ª —Å–æ—Å—Ä–µ–¥–æ—Ç–æ—á–µ–Ω–æ –Ω–∞ –ø–µ—Ä–µ–¥–Ω–µ–º –∫–æ–Ω—Ü–µ —Ç–µ–ª–∞, –∑–∞—Ç–µ–º –Ω–∞ –±—Ä—é—à–Ω–æ–π –∏ –±–æ–∫–æ–≤—ã—Ö –µ–≥–æ —Å—Ç–æ—Ä–æ–Ω–∞—Ö –∏ –º–µ–Ω—å—à–µ –≤—Å–µ–≥–æ –Ω–∞ —Å–ø–∏–Ω–Ω–æ–π –ø–æ–≤–µ—Ä—Ö–Ω–æ—Å—Ç–∏ —Ç–µ–ª–∞ —Ü–µ—Ä–∫–∞—Ä–∏–π. –°–µ–Ω—Å–∏–ª–ª—ã –∏–º–µ—é—Ç—Å—è —Ç–∞–∫–∂–µ –Ω–∞ –ø—Ä–∏—Å–æ—Å–∫–∞—Ö –∏ —Ö–≤–æ—Å—Ç–µ.
–¶–µ—Ä–∫–∞—Ä–∏–∏ –ø–æ –º–µ—Ä–µ —Å–≤–æ–µ–≥–æ —Å–æ–∑—Ä–µ–≤–∞–Ω–∏—è –ø–æ–∫–∏–¥–∞—é—Ç –æ—Ä–≥–∞–Ω–∏–∑–º –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–π –æ—Å–æ–±–∏ (—Ä–µ–¥–∏–∏ –∏ —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç—ã) –∏ –æ–∫–∞–∑—ã–≤–∞—é—Ç—Å—è –≤ –≥–µ–º–æ—Ü–µ–ª–µ –º–æ–ª–ª—é—Å–∫–∞, –≥–¥–µ –æ–Ω–∏ –æ—Å—Ç–∞—é—Ç—Å—è –≤ —Ç–µ—á–µ–Ω–∏–µ –Ω–µ–∫–æ—Ç–æ—Ä–æ–≥–æ –≤—Ä–µ–º–µ–Ω–∏, –ø–æ–∫–∞ –∑–∞–≤–µ—Ä—à–∞–µ—Ç—Å—è –∏—Ö –º–æ—Ä—Ñ–æ–≥–µ–Ω–µ–∑ –∏ –ø—Ä–æ—Ü–µ—Å—Å –Ω–∞–∫–æ–ø–ª–µ–Ω–∏—è –≥–ª–∏–∫–æ–≥–µ–Ω–∞. –£ –Ω–∏–∑—à–∏—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥ (—Å–µ–º. Paramphistomatidae, Notocotylidae) —ç—Ç–æ—Ç –ø—Ä–æ—Ü–µ—Å—Å –¥–ª–∏—Ç–µ–ª—å–Ω—ã–π (7-11 –¥–Ω–µ–π). –ó–∞—Ç–µ–º —Ü–µ—Ä–∫–∞—Ä–∏–∏ –ø—Ä–æ–Ω–∏–∫–∞—é—Ç –≤ –≤–µ–Ω–æ–∑–Ω—ã–µ —Å–æ—Å—É–¥—ã, –≤ —Ä–µ–∫—Ç–∞–ª—å–Ω—ã–π –∏ –ø–æ—á–µ—á–Ω—ã–π —Å–∏–Ω—É—Å—ã, –∫—Ä–æ–≤–µ–Ω–æ—Å–Ω—ã–µ —Å–æ—Å—É–¥—ã –ª–µ–≥–∫–æ–≥–æ, –≤ —Å–µ—Ä–¥—Ü–µ; –ø–æ—Å–ª–µ —á–µ–≥–æ —Ü–µ—Ä–∫–∞—Ä–∏–∏ —Å –∫—Ä–æ–≤—å—é –≤—ã—Ö–æ–¥—è—Ç –≤ –ª–µ–≥–æ—á–Ω—É—é –∞—Ä—Ç–µ—Ä–∏—é, –º–∞–Ω—Ç–∏–π–Ω—ã–µ —Å–∏–Ω—É—Å—ã –∏ –∞–∫—Ç–∏–≤–Ω–æ –≤—ã—Ö–æ–¥—è—Ç –Ω–∞—Ä—É–∂—É.
Заражение второго промежуточного хозяина. Церкарий, подобно мирацидиям, представляет собой расселительную фазу жизненного цикла трематод. Внешняя среда служит для них временной средой обитания, и срок свободной жизни их не продолжителен. Все жизненные функции церкариев подчинены необходимости обеспечить их попадание в организм второго промежуточного хозяина. Для осуществления непосредственного контакта церкариев с хозяином дополнительно выработались специальные адаптации: 1) активные движения в поисках животного хозяина; 2) пассивное парение церкариев в толще воды; 3) «позы ожидания».
Наиболее широкое распространение получили адаптации первого типа ( эхиностоматидные, плеролофоцеркные и стилетные церкарии ); весь период своей свободной жизни эти церкарии непрерывно плавают, не перемещаясь, однако, на большие расстояния. Второй путь приспособлений - парение, не сменяемое фазами активного движения; чаще этот тип адаптации встречается у церкариев, вторым промежуточным хозяином которых служит планктонные морские организмы. Третий тип адаптации заключается в том, что в ходе эволюции у церкариев вырабатывается способность принимать особую «позу», облегчающую возможность быстрого перехода на тело хозяина.
–¶–µ—Ä–∫–∞—Ä–∏–∏, –∞–∫—Ç–∏–≤–Ω–æ –≤–Ω–µ–¥—Ä—è—é—â–∏–µ—Å—è –≤ —Ç–µ–ª–æ —Ö–æ–∑—è–∏–Ω–∞, –ø–æ —Å–ø–æ—Å–æ–±—É —Å–≤–æ–µ–≥–æ –ø—Ä–æ–Ω–∏–∫–Ω–æ–≤–µ–Ω–∏—è –º–æ–≥—É—Ç –±—ã—Ç—å –ø–æ–¥—Ä–∞–∑–¥–µ–ª–µ–Ω—ã –Ω–∞ –¥–≤–µ –≥—Ä—É–ø–ø—ã. –ö –ø–µ—Ä–≤–æ–π –≥—Ä—É–ø–ø–µ –æ—Ç–Ω–æ—Å—è—Ç—Å—è —Å–æ—Å–∞–ª—å—â–∏–∫–∏, —Ü–µ—Ä–∫–∞—Ä–∏–∏ –∫–æ—Ç–æ—Ä—ã—Ö –ø—Ä–æ–Ω–∏–∫–∞—é—Ç –≤ —Ç–µ–ª–æ —Ö–æ–∑—è–∏–Ω–∞, –Ω–µ –Ω–∞—Ä—É—à–∞—è —Ü–µ–ª–æ—Å—Ç–Ω–æ—Å—Ç–∏ –µ–≥–æ –ø–æ–∫—Ä–æ–≤–æ–≤ - —á–µ—Ä–µ–∑ —Ä–æ—Ç–æ–≤–æ–µ, –∞–Ω–∞–ª—å–Ω–æ–µ –∏ –¥—ã—Ö–∞—Ç–µ–ª—å–Ω–æ–µ –æ—Ç–≤–µ—Ä—Å—Ç–∏—è (—Å–µ–º. Echinostomatidae). –¶–µ—Ä–∫–∞—Ä–∏–∏ –≤—Ç–æ—Ä–æ–π –≥—Ä—É–ø–ø—ã –ø—Ä–æ–Ω–∏–∫–∞—é—Ç —á–µ—Ä–µ–∑ –Ω–µ–ø–æ–≤—Ä–µ–∂–¥–µ–Ω–Ω—ã–µ –ø–æ–∫—Ä–æ–≤—ã —Ö–æ–∑—è–∏–Ω–∞. –û–Ω–∏ –º–æ–≥—É—Ç –ø—Ä–æ–±—É—Ä–∞–≤–ª–∏–≤–∞—Ç—å —ç–ø–∏—Ç–µ–ª–∏–π –º–∞–Ω—Ç–∏–∏ –º–æ–ª–ª—é—Å–∫–æ–≤, –∫–æ–∂–Ω–æ-–º—É—Å–∫—É–ª—å–Ω—ã–π –º–µ—à–æ–∫ –ø–∏—è–≤–æ–∫, —Ö–∏—Ç–∏–Ω —á–ª–µ–Ω–∏—Å—Ç–æ–Ω–æ–≥–∏—Ö, –º–Ω–æ–≥–æ—Å–ª–æ–π–Ω—ã–µ –ø–æ–∫—Ä–æ–≤—ã –ø–æ–∑–≤–æ–Ω–æ—á–Ω—ã—Ö –∂–∏–≤–æ—Ç–Ω—ã—Ö.
–ü—Ä–æ—Ü–µ—Å—Å –∏–Ω—Ü–∏—Å—Ç–∏—Ä–æ–≤–∞–Ω–∏—è (—Ü–∏—Å—Ç–æ–≥–æ–Ω–∏—è). –ë–æ–ª—å—à–∞—è —á–∞—Å—Ç—å —Ü–µ—Ä–∫–∞—Ä–∏–µ–≤, –æ—Ç–Ω–æ—Å—è—â–∏—Ö—Å—è –∫ –Ω–∏–∑—à–∏–º —Ç—Ä–µ–º–∞—Ç–æ–¥–∞–º (—Å–µ–º. Fasciolidae, Paramphistomatidae, Notocotylidae –∏ –¥—Ä.), –ø–æ—Å–ª–µ –≤—ã—Ö–æ–¥–∞ –∏–∑ –º–æ–ª–ª—é—Å–∫–∞ –∏–Ω—Ü–∏—Å—Ç–∏—Ä—É—é—Ç—Å—è –≤–æ –≤–Ω–µ—à–Ω–µ–π —Å—Ä–µ–¥–µ –∑–∞ —Å—á–µ—Ç —Å–µ–∫—Ä–µ—Ç–∞, –ø—Ä–æ–¥—É—Ü–∏—Ä—É–µ–º–æ–≥–æ —Ü–∏—Å—Ç–æ–≥–µ–Ω–Ω—ã–º–∏ –∂–µ–ª–µ–∑–∞–º–∏ –∏, –ø—Ä–∏–∫—Ä–µ–ø–ª—è—è—Å—å –∫ —Ä–∞—Å—Ç–µ–Ω–∏—è–º, –ø—Ä–µ–≤—Ä–∞—â–∞—é—Ç—Å—è –≤ –∞–¥–æ–ª–µ—Å–∫–∞—Ä–∏–∏.
–¶–µ—Ä–∫–∞—Ä–∏–∏, –∏–Ω—Ü–∏—Å—Ç–∏—Ä–æ–≤–∞–Ω–∏–µ –∫–æ—Ç–æ—Ä—ã—Ö –ø—Ä–æ–∏—Å—Ö–æ–¥–∏—Ç –≤ –æ—Ä–≥–∞–Ω–∏–∑–º–µ –≤—Ç–æ—Ä–æ–≥–æ –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω–æ–≥–æ —Ö–æ–∑—è–∏–Ω–∞, –æ—Ç–±—Ä–∞—Å—ã–≤–∞—é—Ç —Ö–≤–æ—Å—Ç –≤ –º–æ–º–µ–Ω—Ç –∏—Ö –≤–Ω–µ–¥—Ä–µ–Ω–∏—è –ø–æ–¥ –ø–æ–∫—Ä–æ–≤—ã —Ö–æ–∑—è–∏–Ω–∞. –•–∞—Ä–∞–∫—Ç–µ—Ä —Å—Ç—Ä–æ–µ–Ω–∏—è —Ü–∏—Å—Ç, –∏—Ö –ª–æ–∫–∞–ª–∏–∑–∞—Ü–∏—è –≤ –æ—Ä–≥–∞–Ω–∏–∑–º–µ —Ö–æ–∑—è–∏–Ω–∞, —Å—Ä–æ–∫–∏ –∏–Ω—Ü–∏—Å—Ç–∏—Ä–æ–≤–∞–Ω–∏—è –ø–æ–¥–≤–µ—Ä–≥–Ω—É—Ç—ã –∑–Ω–∞—á–∏—Ç–µ–ª—å–Ω—ã–º –∫–æ–ª–µ–±–∞–Ω–∏—è–º. –ë–∏–æ–ª–æ–≥–∏—á–µ—Å–∫–æ–µ –∑–Ω–∞—á–µ–Ω–∏–µ —Ü–∏—Å—Ç —Å–≤–æ–¥–∏—Ç—Å—è –≤ –æ—Å–Ω–æ–≤–Ω–æ–º –∫ —Ä–æ–ª–∏ –∑–∞—â–∏—Ç–Ω—ã—Ö –æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏–π; —Ü–∏—Å—Ç—ã –∏–∑–æ–ª–∏—Ä—É—é—Ç –º–µ—Ç–∞—Ü–µ—Ä–∫–∞—Ä–∏–π –æ—Ç –Ω–µ–±–ª–∞–≥–æ–ø—Ä–∏—è—Ç–Ω—ã—Ö –≤–æ–∑–¥–µ–π—Å—Ç–≤–∏–π –≤–Ω–µ—à–Ω–µ–π —Å—Ä–µ–¥—ã –∏–ª–∏ –∏–º–º—É–Ω–æ–ª–æ–≥–∏—á–µ—Å–∫–∏—Ö —Ä–µ–∞–∫—Ü–∏–π —Ö–æ–∑—è–∏–Ω–∞, –Ω–æ –æ–±–æ–ª–æ—á–∫–∞ —Ü–∏—Å—Ç –ø–æ–ª—É–ø—Ä–æ–Ω–∏—Ü–∞–µ–º–∞, –æ–Ω–∞ –æ–±–µ—Å–ø–µ—á–∏–≤–∞–µ—Ç –¥–æ—Å—Ç—É–ø –ø–∏—â–µ–≤—ã—Ö –≤–µ—â–µ—Å—Ç–≤, –Ω–µ–æ–±—Ö–æ–¥–∏–º—ã—Ö –º–µ—Ç–∞—Ü–µ—Ä–∫–∞—Ä–∏—è–º.
–ú–µ—Ç–∞—Ü–µ—Ä–∫–∞—Ä–∏–∏ —Ç—Ä–µ–º–∞—Ç–æ–¥ –∏–Ω—Ü–∏—Å—Ç–∏—Ä—É—é—Ç—Å—è –≤ –ø–æ–ª–æ—Å—Ç—è—Ö —Ç–µ–ª–∞ –º–æ–ª–ª—é—Å–∫–æ–≤, —Ä—ã–±, –∞–º—Ñ–∏–±–∏–π, —Å–æ–æ–±—â–∞—é—â–∏—Ö—Å—è —Å –≤–Ω–µ—à–Ω–µ–π —Å—Ä–µ–¥–æ–π (—Å–µ–º. Echinostomatidae); –≤ —Ç–æ–ª—â–µ –º–∞–Ω—Ç–∏–∏ –º–æ–ª–ª—é—Å–∫–æ–≤, –≤ –ø–∞—Ä–µ–Ω—Ö–∏–º–µ –ø–∏—è–≤–æ–∫, –≤ —Å–æ–±—Å—Ç–≤–µ–Ω–Ω–æ–π –∫–æ–∂–µ –∏ –ø–æ–¥–∫–æ–∂–Ω–æ–π –∫–ª–µ—Ç—á–∞—Ç–∫–µ —Ä—ã–± –∏ –∞–º—Ñ–∏–±–∏–π (—Å–µ–º. Strigeidae, Diplostomatidae).
–ú–µ—Ç–∞—Ü–µ—Ä–∫–∞—Ä–∏—è. –¶–µ—Ä–∫–∞—Ä–∏–∏ –ø–æ—Å–ª–µ –∏–Ω—Ü–∏—Å—Ç–∏—Ä–æ–≤–∞–Ω–∏—è –≤ –æ—Ä–≥–∞–Ω–∏–∑–º–µ –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω–æ–≥–æ —Ö–æ–∑—è–∏–Ω–∞, –ø—Ä–µ—Ç–µ—Ä–ø–µ–≤ –º–µ—Ç–∞–º–æ—Ä—Ñ–æ–∑ –∏–ª–∏ –æ–ø—Ä–µ–¥–µ–ª–µ–Ω–Ω—ã–µ —Å—Ç–∞–¥–∏–∏ –º–æ—Ä—Ñ–æ–≥–µ–Ω–µ–∑–∞, –ø—Ä–µ–≤—Ä–∞—â–∞—é—Ç—Å—è –≤ —Å–ª–µ–¥—É—é—â—É—é —Ñ–∞–∑—É —Ä–∞–∑–≤–∏—Ç–∏—è (–º–µ—Ç–∞—Ü–µ—Ä–∫–∞—Ä–∏—è). –ú–µ—Ç–∞—Ü–µ—Ä–∫–∞—Ä–∏–∏ –≤ —Å—É—â–Ω–æ—Å—Ç–∏ –ø—Ä–µ–¥—Å—Ç–∞–≤–ª—è—é—Ç —Å–æ–±–æ–π –Ω–µ–¥–æ—Ä–∞–∑–≤–∏—Ç–∏—é –º–∞—Ä–∏—Ç—É –∏ —É–∂–µ —Ö–∞—Ä–∞–∫—Ç–µ—Ä–∏–∑—É—é—Ç—Å—è –º–Ω–æ–≥–∏–º–∏ —á–µ—Ä—Ç–∞–º–∏ —Å—Ç—Ä–æ–µ–Ω–∏—è, —Å–≤–æ–π—Å—Ç–≤–µ–Ω–Ω—ã–º–∏ –¥–∞–Ω–Ω–æ–º—É –≤–∏–¥—É.
–°–ò–°–¢–ï–ú–ê–¢–ò–ö–ê –¢–Ý–ï–ú–ê–¢–û–î.
–ö–ª–∞—Å—Å–∏—Ñ–∏–∫–∞—Ü–∏—è —Ç—Ä–µ–º–∞—Ç–æ–¥ –Ω–∞—á–∞–ª–∞—Å—å —Å –ö.–õ–∏–Ω–Ω–µ—è (1758), –ø—Ä–∏–∑–Ω–∞–≤–∞–≤—à–µ–≥–æ –æ–¥–∏–Ω —Ä–æ–¥ Fasciola —Å –¥–≤—É–º—è –≤–∏–¥–∞–º–∏. –í –ø–æ—Å–ª–µ–¥—É—é—â–µ–º –±—ã–ª–∏ –ø—Ä–µ–¥–ø—Ä–∏–Ω—è—Ç—ã –ø–æ–ø—ã—Ç–∫–∏ —Å–∏—Å—Ç–µ–º–∞—Ç–∏–∫–∏ —Ç—Ä–µ–º–∞—Ç–æ–¥ –Ω–∞ –æ—Å–Ω–æ–≤–µ –º–æ—Ä—Ñ–æ–ª–æ–≥–∏–∏ –ø–æ–ª–æ–≤–æ–∑—Ä–µ–ª—ã—Ö —Ç—Ä–µ–º–∞—Ç–æ–¥, —Å—Ç—Ä–æ–µ–Ω–∏—è —ç–∫—Å–∫—Ä–µ—Ç–æ—Ä–Ω–æ–π —Å–∏—Å—Ç–µ–º—ã –∏ –≤–Ω–µ—à–Ω–∏—Ö –ø—Ä–∏–∑–Ω–∞–∫–æ–≤ —Ü–µ—Ä–∫–∞—Ä–∏–µ–≤ –∏ –±–∏–æ–ª–æ–≥–∏–∏ –ø–∞—Ä–∞–∑–∏—Ç–æ–≤. –ó–Ω–∞—á–∏—Ç–µ–ª—å–Ω—ã–π –≤–∫–ª–∞–¥ –≤ —Å–∏—Å—Ç–µ–º–∞—Ç–∏–∫—É —Ç—Ä–µ–º–∞—Ç–æ–¥ –≤–Ω–µ—Å–ª–∏ –≥–µ–ª—å–º–∏–Ω—Ç–æ–ª–æ–≥–∏ –ö.–ò.–°–∫—Ä—è–±–∏–Ω –∏ –Ý.–°.–®—É–ª—å—Ü. –ù–∏–∂–µ –ø—Ä–∏–≤–æ–¥–∏—Ç—Å—è —Å–∏—Å—Ç–µ–º–∞—Ç–∏–∫–∞ —Ç—Ä–µ–º–∞—Ç–æ–¥, –ø—Ä–∏–Ω—è—Ç–∞—è –≤ –±–æ–ª—å—à–∏–Ω—Å—Ç–≤–µ —Å—Ç—Ä–∞–Ω.
–ö–õ–ê–°–° Trematoda Rudolphi, 1808.
1.ПОДКЛАСС Bucephalidae – ротовое отверстие на брюшной стороне тела, около его середине, лишено ротовой присоски. Кишечник простой, мешковидный, короткий. На переднем конце имеется мышечная присоска или хоботок со щупальцами. Подкласс включает один отряд и одно семейство. Паразиты карповых, окуневых, лососевых, тресковых (налим) рыб и амфибий.
–û–¢–Ý–Ø–î Bucephalida
–°–µ–º. Bucephalidea Poche, 1907.
–°–µ–º–µ–π—Å—Ç–≤–æ –æ–±—ä–µ–¥–∏–Ω—è–µ—Ç –Ω–µ—Å–∫–æ–ª—å–∫–æ —Ä–æ–¥–æ–≤ –∏ –±–æ–ª–µ–µ 100 –≤–∏–¥–æ–≤.
2.–ü–û–î–ö–õ–ê–°–° Prosostomidea Skrjabin et Gusochanskaja, 1962 ‚Äì –æ—Ä–≥–∞–Ω—ã –ø—Ä–∏–∫—Ä–µ–ø–ª–µ–Ω–∏—è –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω—ã —Ä–∞–∑–ª–∏—á–Ω–æ —Ä–∞–∑–≤–∏—Ç—ã–º–∏ —Ä–æ—Ç–æ–≤–æ–π –∏ –±—Ä—é—à–Ω–æ–π –ø—Ä–∏—Å–æ—Å–∫–∞–º–∏, –æ–¥–Ω–∞ –∏–∑ –Ω–∏—Ö –∏–ª–∏ –æ–±–µ –º–æ–≥—É—Ç –æ—Ç—Å—É—Ç—Å—Ç–≤–æ–≤–∞—Ç—å. –Ý–∞–∑–≤–∏—Ç–∏–µ —Å–ª–æ–∂–Ω–æ–µ: —Å —á–µ—Ä–µ–¥–æ–≤–∞–Ω–∏–µ–º –ø–æ–∫–æ–ª–µ–Ω–∏–π –∏ —Å–º–µ–Ω–æ–π —Ö–æ–∑—è–µ–≤. –ü–æ–¥–∫–ª–∞—Å—Å —Å–æ—Å—Ç–æ–∏—Ç –∏–∑ —Ç—Ä–µ—Ö –æ—Ç—Ä—è–¥–æ–≤.
1. –û–¢–Ý–Ø–î Fasciolida, Skrjabin et Guschanskaja, 1962, –≤–∫–ª—é—á–∞–µ—Ç 12 –ø–æ–¥–æ—Ç—Ä—è–¥–æ–≤:
–∞) –ü–û–î–û–¢–Ý–Ø–î Schistosomatata Skrjabin et Schulz, 1937
- —Ä–∞–∑–¥–µ–ª—å–Ω–æ–ø–æ–ª—ã–µ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã, –ø—Ä–∏—Å–æ—Å–∫–∏ —Ä–∞–∑–≤–∏—Ç—ã —Å–ª–∞–±–æ –∏–ª–∏ –≤–æ–≤—Å–µ –æ—Ç—Å—É—Ç—Å—Ç–≤—É—é—Ç; —è–π—Ü–∞ –æ–±—ã—á–Ω–æ —Å —à–∏–ø–∏–∫–∞–º–∏. –ü–∞—Ä–∞–∑–∏—Ç–∏—Ä—É—é—Ç –≤ –∫—Ä–æ–≤–µ–Ω–æ—Å–Ω–æ–π —Å–∏—Å—Ç–µ–º–µ –ø—Ç–∏—Ü –∏ –º–ª–µ–∫–æ–ø–∏—Ç–∞—é—â–∏—Ö;
–±) –ü–û–î–û–¢–Ý–Ø–î Sanquinicolata Skrjabin et Schulz ,1937
–≥–µ—Ä–º–∞—Ñ—Ä–æ–¥–∏—Ç–Ω—ã–µ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã, –ø–∞—Ä–∞–∑–∏—Ç—ã –∫—Ä–æ–≤–∏ —Ä—ã–± –∏ —Ä–µ–ø—Ç–∏–ª–∏–π. –ü—Ä–∏—Å–æ—Å–∫–∏ –∏–º–µ—é—Ç—Å—è –∏–ª–∏ –æ—Ç—Å—É—Ç—Å—Ç–≤—É—é—Ç, —Ñ–∞—Ä–∏–Ω–∫—Å–∞ –Ω–µ—Ç, –º–∞—Ç–∫–∞ –∫–æ—Ä–æ—Ç–∫–∞—è. –í–∫–ª—é—á–∞–µ—Ç —Å–µ–º. Spirorchidae –∏ Sanquinicolidae. –ü–æ—Å–ª–µ–¥–Ω–∏–µ –º–µ–ª–∫–∏–µ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã (2 –º–º –¥–ª–∏–Ω—ã), –º–∞—Ç–∫–∞ —É –Ω–∏—Ö –æ—Ç—Å—É—Ç—Å—Ç–≤—É–µ—Ç, –≤ –æ–æ—Ç–∏–ø–µ –∏–º–µ–µ—Ç—Å—è —Ç–æ–ª—å–∫–æ –æ–¥–Ω–æ —è–π—Ü–æ, –∏–Ω–æ–≥–¥–∞ –∏–º–µ–µ—Ç—Å—è —É–∫–æ—Ä–æ—á–µ–Ω–Ω–∞—è –º–∞—Ç–∫–∞. –í —Ä—ã–±–∞—Ö –ø—Ä–µ—Å–Ω—ã—Ö –≤–æ–¥–æ–µ–º–æ–≤ –Ý–§ –≤—Å—Ç—Ä–µ—á–∞—é—Ç—Å—è –≤–æ—Å–µ–º—å –≤–∏–¥–æ–≤ –∏–∑ —Ä–æ–¥–∞ Sanquinicola, –Ω–æ –Ω–∞–∏–±–æ–ª–µ–µ –ø–∞—Ç–æ–≥–µ–Ω–µ–Ω S.inermis;
–≤) –ü–û–î–û–¢–Ý–Ø–î Pronocephalata Skrjabin ,1955 ‚Äì –±—Ä—é—à–Ω–∞—è –ø—Ä–∏—Å–æ—Å–∫–∞ –∏ —Ñ–∞—Ä–∏–Ω–∫—Å –æ—Ç—Å—É—Ç—Å—Ç–≤—É—é—Ç, –ø–æ–ª–æ–≤—ã–µ –∂–µ–ª–µ–∑—ã —Ä–∞—Å–ø–æ–ª–æ–∂–µ–Ω—ã –≤ –∑–∞–¥–Ω–µ–π —á–∞—Å—Ç–∏ —Ç–µ–ª–∞. –ü–∞—Ä–∞–∑–∏—Ç—ã –ø–∏—â–µ–≤–∞—Ä–∏—Ç–µ–ª—å–Ω–æ–≥–æ —Ç—Ä–∞–∫—Ç–∞ –∏ –æ—Ä–≥–∞–Ω–æ–≤ –¥—ã—Ö–∞–Ω–∏—è –ø–æ–∑–≤–æ–Ω–æ—á–Ω—ã—Ö. –ù–∞–∏–±–æ–ª–µ–µ —á–∞—Å—Ç–æ –≤—Å—Ç—Ä–µ—á–∞—é—Ç—Å—è –≤–∏–¥—ã –∏–∑ —Å–µ–º–µ–π—Å—Ç–≤:
—Å–µ–º. Notocotylidae Luhe, 1909 ‚Äì –º–µ–ª–∫–∏–µ –∏ —Å—Ä–µ–¥–Ω–µ–π –≤–µ–ª–∏—á–∏–Ω—ã —Ç—Ä–µ–º–∞—Ç–æ–¥—ã (3-5 –º–º –¥–ª–∏–Ω—ã), —Ç–µ–ª–æ –≤ –∑–∞–¥–Ω–µ–π —á–∞—Å—Ç–∏ –≥—É—Å—Ç–æ —É—Å–µ—è–Ω–æ –º–µ–ª—å—á–∞–π—à–∏–º–∏ —à–∏–ø–∏–∫–∞–º–∏. –Ý–æ—Ç–æ–≤–∞—è –ø—Ä–∏—Å–æ—Å–∫–∞ –ø—Ä–æ—Å—Ç–∞—è, –ø–∏—â–µ–≤–æ–¥ –∫–æ—Ä–æ—Ç–∫–∏–π, —Ñ–∞—Ä–∏–Ω–∫—Å –æ—Ç—Å—É—Ç—Å—Ç–≤—É–µ—Ç. –°–µ–º–µ–π—Å—Ç–≤–æ —Å–æ–¥–µ—Ä–∂–∏—Ç 9 —Ä–æ–¥–æ–≤; —É –¥–æ–º–∞—à–Ω–∏—Ö –≤–æ–¥–æ–ø–ª–∞–≤–∞—é—â–∏—Ö –ø—Ç–∏—Ü –ø–∞—Ä–∞–∑–∏—Ç–∏—Ä—É—é—Ç –ø—Ä–µ–¥—Å—Ç–∞–≤–∏—Ç–µ–ª–∏ —Ä–æ–¥–æ–≤ Notocotylus (N. –∞ttenuatus, N. chionis), Catatropis (–°. verrucosa).
сем. Pronocephalidae Looss, 1902 – мелкие и средние трематоды, передний конец снабжен воротничком, на заднем – два выступа; яйца многочисленные, с филаментами. Паразитируют в кишечнике морских и пресноводных черепах;
–≥) –ü–û–î–û–¢–Ý–Ø–î Cyclocoeliata La Rue, 1957 ‚Äì –∫—Ä—É–ø–Ω—ã—Ö –∏ –º–µ–ª–∫–∏—Ö —Ä–∞–∑–º–µ—Ä–æ–≤ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã, –∫—É—Ç–∏–∫—É–ª–∞ –±–µ–∑ —à–∏–ø–æ–≤, —Ä–æ—Ç–æ–≤–∞—è –ø—Ä–∏—Å–æ—Å–∫–∞ –æ—Ç—Å—É—Ç—Å—Ç–≤—É–µ—Ç –±—Ä—é—à–Ω–∞—è –ø—Ä–∏—Å–æ—Å–∫–∞ —Ä—É–¥–∏–º–µ–Ω—Ç–∞—Ä–Ω–∞. –ó—Ä–µ–ª—ã–µ —è–π—Ü–∞ –≤ –º–∞—Ç–∫–µ —Å–æ–¥–µ—Ä–∂–∞—Ç –º–∏—Ä–∞—Ü–∏–¥–∏–µ–≤, —Å–Ω–∞–±–∂–µ–Ω–Ω—ã—Ö –ø–∏–≥–º–µ–Ω—Ç–Ω—ã–º –≥–ª–∞–∑–∫–æ–º; –º–∏—Ä–∞—Ü–∏–¥–∏–∏ –º–æ–≥—É—Ç –≤—ã—Ö–æ–¥–∏—Ç—å –∏–∑ —Å–∫–æ—Ä–ª—É–ø—ã –µ—â–µ –≤–Ω—É—Ç—Ä–∏ –º–∞—Ç–∫–∏. –ü–∞—Ä–∞–∑–∏—Ç–∏—Ä—É—é—Ç –≤ –ø–æ–ª–æ—Å—Ç–∏ —Ç–µ–ª–∞, –Ω–æ—Å–æ–≤—ã—Ö –∏ –≥–ª–∞–∑–Ω—ã—Ö –ø–æ–ª–æ—Å—Ç—è—Ö, –≤ –≤–æ–∑–¥—É—Ö–æ–Ω–æ—Å–Ω—ã—Ö –º–µ—à–∫–∞—Ö –ø—Ä–∏–≤–æ–¥–Ω—ã—Ö –ø—Ç–∏—Ü, –ø—Ä–µ–∏–º—É—â–µ—Å—Ç–≤–µ–Ω–Ω–æ –¥–∏–∫–∏—Ö(Tracheophilus sisowi);
–¥) –ü–û–î–û–¢–Ý–Ø–î Echinostomatata Szidat, 1936 - –∏–º–µ—é—Ç –≤–æ–æ—Ä—É–∂–µ–Ω–Ω—ã–π –∏–ª–∏ –Ω–µ–≤–æ–æ—Ä—É–∂–µ–Ω–Ω—ã–π –∞–¥–æ—Ä–∞–ª—å–Ω—ã–π –¥–∏—Å–∫ (–≤–æ—Ä–æ—Ç–Ω–∏—á–æ–∫):
сем. Echinostomatidae Dietz, 1909 – есть адоральный диск, снабженный шипиками. Кутикула в передней части имеет мелкие шипики, брюшная присоска крупная. Паразитируют в кишечнике, желчных ходах и фабрициевой сумке птиц. Это семейство содержит 40 родов, объединяющих около 300 видов. Наиболее богаты видами роды Echinostoma (E.revolutum, E.miyagawai , E.paraulum), Echinoparyphium (E. recurvatum), Echinochasmus (E.beleocephalus).
сем. Cotylotretidae Skrjabin at Baschkirova, 1956 – адоральный диск отсутствует, головной конец лишен шипов. Паразитируют у птиц, грызунов и ластоногих.
–µ) –ü–û–î–û–¢–Ý–Ø–î Heterophyata Morosow, 1955 - –º–µ–ª–∫–∏–µ –∏ —Å—Ä–µ–¥–Ω–µ–π –≤–µ–ª–∏—á–∏–Ω—ã —Ç—Ä–µ–º–∞—Ç–æ–¥—ã, —Å–Ω–∞–±–∂–µ–Ω—ã —Ä–æ—Ç–æ–≤–æ–π –∏ –±—Ä—é—à–Ω–æ–π –ø—Ä–∏—Å–æ—Å–∫–∞–º–∏. –¶–µ—Ä–∫–∞—Ä–∏–∏ –ª–∏—à–µ–Ω—ã –ø—Ä–∏—Å–æ—Å–∫–∏, –º–µ—Ç–∞—Ü–µ—Ä–∫–∞—Ä–∏–∏ ‚Äì –≤ —Ä—ã–±–∞—Ö –∏–ª–∏ —Ä–∞–∫–æ–æ–±—Ä–∞–∑–Ω—ã—Ö, –ø–æ–ª–æ–≤–æ–∑—Ä–µ–ª—ã–µ –ø–∞—Ä–∞–∑–∏—Ç—ã ‚Äì –≤ –ø–∏—â–µ–≤–∞—Ä–∏—Ç–µ–ª—å–Ω–æ–º —Ç—Ä–∞–∫—Ç–µ, –∂–µ–ª—á–Ω—ã—Ö —Ö–æ–¥–∞—Ö –ø–µ—á–µ–Ω–∏, –∂–µ–ª—á–Ω–æ–º –ø—É–∑—ã—Ä–µ –º–ª–µ–∫–æ–ø–∏—Ç–∞—é—â–∏—Ö.
–°–µ–º. Heterophyidae Odhner, 1914 ‚Äì –º–µ–ª–∫–∏–µ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã (2,5 –º–º), –∫—É—Ç–∏–∫—É–ª–∞ –ø–æ–∫—Ä—ã—Ç–∞ —à–∏–ø–∞–º–∏. –Ý–∞–∑–≤–∏–≤–∞—é—Ç—Å—è —Å —É—á–∞—Å—Ç–∏–µ–º –¥–≤—É—Ö –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω—ã—Ö —Ö–æ–∑—è–µ–≤ (–º–æ–ª–ª—é—Å–∫–∏ –∏ —Ä—ã–±—ã). –ü–∞—Ä–∞–∑–∏—Ç–∏—Ä—É—é—Ç –≤ –∫–∏—à–µ—á–Ω–∏–∫–µ —á–µ–ª–æ–≤–µ–∫–∞.
Сем. Galactosomidae Morosow, 1950 – 3–3,5 мм длины, брюшная присоска маленькая. Паразитируют в кишечнике птиц и млекопитающих. Метацеркарии локализуются в чешуе, жабрах, плавниках и других органах рыб.
Сем. Opisthorchidae Braun, 1901 – средней величины трематоды; присоски сближены; брюшная присоска слабо развита. Яйца мелкие, коричневого цвета. Семейство объединяет более 20 родов (80 видов). Наибольшее значение имеют роды: Opisthorchis (O.felineus), Clonorchis (C.sinensis) – паразиты печени у человека.
–∂) –ü–û–î–û–¢–Ý–Ø–î Azygiata La Rue, 1957 - —Ç—Ä–µ–º–∞—Ç–æ–¥—ã –∏–º–µ—é—Ç —Ä–æ—Ç–æ–≤—É—é –∏ –±—Ä—é—à–Ω—É—é –ø—Ä–∏—Å–æ—Å–∫–∏, –º—ã—à–µ—á–Ω—ã–π —Ñ–∞—Ä–∏–Ω–∫—Å. –ü–æ–ª–æ–≤–æ–∑—Ä–µ–ª—ã–µ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã –ø–∞—Ä–∞–∑–∏—Ç–∏—Ä—É—é—Ç –≤ –∫–∏—à–µ—á–Ω–∏–∫–µ —Ä—ã–± (–æ–∫—É–Ω–µ–π, —â—É–∫);
–∑) –ü–û–î–û–¢–Ý–Ø–î Allocreadiata Skrjabin, Petrow et Kovae 1958 - –æ–≤–∞–ª—å–Ω—ã–µ –∏–ª–∏ —É–¥–ª–∏–Ω–µ–Ω–Ω—ã–µ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã —Å —Ö–æ—Ä–æ—à–æ —Ä–∞–∑–≤–∏—Ç—ã–º –ø—Ä–∏—Å–æ—Å–∫–∞–º–∏. –Ø–π—Ü–∞ –∫—Ä—É–ø–Ω—ã–µ, —Å –∫—Ä—ã—à–µ—á–∫–∞–º–∏. –ü–∞—Ä–∞–∑–∏—Ç—ã –ø–∏—â–µ–≤–∞—Ä–∏—Ç–µ–ª—å–Ω–æ–≥–æ —Ç—Ä–∞–∫—Ç–∞ —Å—É–¥–∞–∫–æ–≤, –æ–∫—É–Ω–µ–π, —â—É–∫, –ª–æ—Å–æ—Å–µ–π, —Ö–∞—Ä–∏—É—Å–æ–≤, –Ω–∞–ª–∏–º–æ–≤ (—Å–µ–º. Bunoderidae Nicoll, 1914 ‚Äì Bunodera Euciopercae, Crepidostomum farionis) –ø—Ä–µ—Å–Ω–æ–≤–æ–¥–Ω—ã—Ö –∏ –º–æ—Ä—Å–∫–∏—Ö —Ä—ã–± (—Å–µ–º. Opecoelidae Ozaki 1925 );
–∏) –ü–û–î–û–¢–Ý–Ø–î Fasciolata Skrjabin et Schulz, 1935 ‚Äì —Ç—Ä–µ–º–∞—Ç–æ–¥—ã —Å –æ—á–µ–Ω—å —Ä–∞–∑–Ω–æ–æ–±—Ä–∞–∑–Ω–æ–π —Ñ–æ—Ä–º–æ–π —Ç–µ–ª–∞, —Ä–∞–∑–º–µ—Ä–æ–º –æ—Ç 1 –º–º –¥–æ –Ω–µ—Å–∫–æ–ª—å–∫–∏—Ö —Å–º. –ö —ç—Ç–æ–º—É –æ—Ç—Ä—è–¥—É –æ—Ç–Ω–æ—Å—è—Ç—Å—è –±–æ–ª–µ–µ 40 —Å–µ–º–µ–π—Å—Ç–≤. –ù–∞–∏–±–æ–ª—å—à–µ–µ –∑–Ω–∞—á–µ–Ω–∏–µ –∏–º–µ—é—Ç —Å–µ–º–µ–π—Å—Ç–≤–∞:
–°–µ–º. Fasciolidae Railliet, 1895 ‚Äì –∫—Ä—É–ø–Ω—ã–µ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã —Å –±–ª–∏–∑–∫–æ —Ä–∞—Å–ø–æ–ª–æ–∂–µ–Ω–Ω—ã–º–∏ —Ä–æ—Ç–æ–≤–æ–π –∏ –±—Ä—é—à–Ω–æ–π –ø—Ä–∏—Å–æ—Å–∫–∞–º–∏, —Å–µ–º—è–ø—Ä–∏–µ–º–Ω–∏–∫ –æ—Ç—Å—É—Ç—Å—Ç–≤—É–µ—Ç. –°–µ–º–µ–π—Å—Ç–≤–æ –≤–∫–ª—é—á–∞–µ—Ç –¥–æ 10 –≤–∏–¥–æ–≤ –ø—è—Ç–∏ —Ä–æ–¥–æ–≤, –ø–∞—Ä–∞–∑–∏—Ç–∏—Ä—É—é—â–∏—Ö –≤ –∂–µ–ª—á–Ω—ã—Ö —Ö–æ–¥–∞—Ö –ø–µ—á–µ–Ω–∏ (—Ä–µ–∂–µ –≤ –∫–∏—à–µ—á–Ω–∏–∫–µ) –Ω–∞–∑–µ–º–Ω—ã—Ö –º–ª–µ–∫–æ–ø–∏—Ç–∞—é—â–∏—Ö (–∫–æ–ø—ã—Ç–Ω—ã—Ö), —Ç–∞–∫–∂–µ —á–µ–ª–æ–≤–µ–∫–∞. –®–∏—Ä–æ–∫–æ —Ä–∞—Å–ø—Ä–æ—Å—Ç—Ä–∞–Ω–µ–Ω—ã –∏ –Ω–∞–∏–±–æ–ª–µ–µ –ø–∞—Ç–æ–≥–µ–Ω–Ω—ã —Å–ª–µ–¥—É—é—â–∏–µ –≤–∏–¥—ã: Fasciola hepatica, E.gigantica (–≤ –°–ù–ì), F.californisa, F.halli –≤ (–°–®–ê). –Ý–∞–∑–≤–∏–≤–∞—é—Ç—Å—è —Å –æ–¥–Ω–∏–º –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω—ã–º —Ö–æ–∑—è–∏–Ω–æ–º ‚Äì –º–æ–ª–ª—é—Å–∫–∞–º–∏. –ê–¥–æ–ª–µ—Å–∫–∞—Ä–∏–∏ –∏–Ω—Ü–∏—Å—Ç–∏—Ä—É—é—Ç—Å—è –Ω–∞ –≤–æ–¥–Ω—ã—Ö –∏ –ø—Ä–∏–±—Ä–µ–∂–Ω—ã—Ö —Ä–∞—Å—Ç–µ–Ω–∏—è—Ö –∏–ª–∏ –Ω–∞ –ø–æ–≤–µ—Ä—Ö–Ω–æ—Å—Ç–∏ –≤–æ–¥—ã.
Сем. Dicrocoeliidae Odhner, 1911 – удлиненные либо круглые трематоды. Присоски сближены, лежат в передней части тела, фаринкс развит хорошо, имеется пищевод. Хорошо развита половая бурса, содержит невооруженный циррус, семенной пузырь.
–í–∏–¥ Dicrocoelium lanceatum ‚Äì –ª–∞–Ω—Ü–µ—Ç–æ–≤–∏–¥–Ω–∞—è –¥–≤—É—É—Å—Ç–∫–∞ ‚Äì –ø–∞—Ä–∞–∑–∏—Ç –∂–µ–ª—á–Ω—ã—Ö –ø—Ä–æ—Ç–æ–∫–æ–≤ –∏ –∂–µ–ª—á–Ω–æ–≥–æ –ø—É–∑—ã—Ä—è –∫–æ–ø—ã—Ç–Ω—ã—Ö, –∑–∞–π—Ü–µ–æ–±—Ä–∞–∑–Ω—ã—Ö –∏ –≥—Ä—ã–∑—É–Ω–æ–≤. –Ý–∞–∑–≤–∏—Ç–∏–µ –ø—Ä–æ–∏—Å—Ö–æ–¥–∏—Ç —Å–æ —Å–º–µ–Ω–æ–π –¥–≤—É—Ö –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω—ã—Ö —Ö–æ–∑—è–µ–≤ ‚Äì –Ω–∞–∑–µ–º–Ω—ã—Ö –º–æ–ª–ª—é—Å–∫–æ–≤ –∏ –Ω–∞—Å–µ–∫–æ–º—ã—Ö (–º—É—Ä–∞–≤—å–µ–≤).
Вид Eurytrema pancreaticum – паразитирует в протоках поджелудочной железы у крупного и мелкого рогатого скота.
Сем. Plagiorchidae Luhe, 1901 – мелкие трематоды, кутикула их покрыта шипиками. Половое отверстие внутри брюшной присоски, семенники – позади яичника, половая бурса длинная, заходит за уровень брюшной присоски, внутри содержит семенной пузырек. Циррус короткий или длинный. Матка располагается позади брюшной присоски, проходит между семенниками, достигая заднего конца тела. Яйца мелкие, церкарии имеют стилет. Семейство включает много родов, но наибольшее значение имеет род Plagiorchis, вид P.orcuatus – паразит яйцевода и фабрициевой сумки домашних кур. P. maculosus – диких птиц, P.cutamiatis – ондатры.
Сем. Prostogonimidae Nicoll, - 1924 – мелких и средних размеров трематоды, с суженным передним и расширенным задним концом тела. Кутикула покрыта шипиками. Паразиты яйцевода, фабрициевой сумки птиц. Паразиты развиваются с двумя промежуточными хозяевами – моллюсками и насекомыми. Типичный род - Prosthogonimus; наибольшее ветеринарное значение имеют P.ovatus P. cuneatus P.macrorchis;
–∫) –ü–û–î–û–¢–Ý–Ø–î Paramphistomatata Szidat, 1936 Skrjabin
et Schulz, 1937 - –∏–º–µ–µ—Ç —Å–¥–≤–∏–Ω—É—Ç—É—é –∫ –∑–∞–¥–Ω–µ–º—É –∫–æ–Ω—Ü—É —Ç–µ–ª–∞ –±—Ä—é—à–Ω—É—é –ø—Ä–∏—Å–æ—Å–∫—É, –∫–∏—à–µ—á–Ω—ã–µ —Å—Ç–≤–æ–ª—ã –º–æ—â–Ω–æ —Ä–∞–∑–≤–∏—Ç—ã. –Ý–∞–∑–≤–∏–≤–∞—é—Ç—Å—è –ø—Ä–∏ —É—á–∞—Å—Ç–∏–∏ –æ–¥–Ω–æ–≥–æ –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω–æ–≥–æ —Ö–æ–∑—è–∏–Ω–∞. –¶–µ—Ä–∫–∞—Ä–∏–∏ –≤–æ –≤–Ω–µ—à–Ω–µ–π —Å—Ä–µ–¥–µ –ø—Ä–µ–≤—Ä–∞—â–∞—é—Ç—Å—è –≤ –∞–¥–æ–ª–µ—Å–∫–∞—Ä–∏–µ–≤. –ü–∞—Ä–∞–∑–∏—Ç—ã —Ä—ã–±, –∞–º—Ñ–∏–±–∏–π, –ø—Ç–∏—Ü –∏ –º–ª–µ–∫–æ–ø–∏—Ç–∞—é—â–∏—Ö. –ü–æ–¥–æ—Ç—Ä—è–¥ –≤–∫–ª—é—á–∞–µ—Ç –¥–µ—Å—è—Ç—å —Å–µ–º–µ–π—Å—Ç–≤, –Ω–∞–∏–±–æ–ª—å—à–µ–µ –≤–µ—Ç–µ—Ä–∏–Ω–∞—Ä–Ω–æ–µ –∑–Ω–∞—á–µ–Ω–∏–µ –∏–º–µ—é—Ç —Å–µ–º–µ–π—Å—Ç–≤–∞:
Сем. Paramphistomatidae Fischceder, 1901 – тело коническое, цилиндрическое (2-12 мм), кутикула без шипиков. Задняя присоска лежит апикально, реже вентрально. Половая бурса отсутствует. Паразитирует в желудке, рубце и желчных ходах копытных млекопитающих. В это семейство входят девять родов, объединяющих около 60 видов. У копытных в СНГ зарегистрировано несколько родов Paramphistomum, Calicophoron,Liorchis.
–°–µ–º.Gastrothylacidae Stiles et Goldberger, 1910 —Ä–∞—Å–ø—Ä–æ—Å—Ç—Ä–∞–Ω–µ–Ω—ã –≤ —Å—Ç—Ä–∞–Ω–∞—Ö —Å –∂–∞—Ä–∫–∏–º –∫–ª–∏–º–∞—Ç–æ–º, –æ–ø–∏—Å–∞–Ω–æ 20 –≤–∏–¥–æ–≤, –≤ –Ω–∞—à–µ–π —Å—Ç—Ä–∞–Ω–µ –≤—Å—Ç—Ä–µ—á–∞—é—Ç—Å—è 2 –≤–∏–¥–∞: Gastrothyla—Ö crymenifer (1847) –∏ Fischoederius elongatus (1883).
2. –û–¢–Ý–Ø–î Strigeidida La Rue, 1926 (Sudaricov, 1959) ‚Äì –∏–º–µ—é—Ç –ø–æ–∑–∞–¥–∏ –±—Ä—é—à–Ω–æ–π –ø—Ä–∏—Å–æ—Å–∫–∏ –æ—Ä–≥–∞–Ω –ë—Ä–∞–Ω–¥–µ—Å–∞ ‚Äì —Å–ª–æ–∂–Ω—ã–π —Å–µ–∫—Ä–µ—Ç–æ—Ä–Ω–æ - —Ñ–∏–∫—Å–∞—Ç–æ—Ä–Ω—ã–π –∞–ø–ø–∞—Ä–∞—Ç; –ø–æ–ª–æ–≤–æ–µ –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ –Ω–∞—Ö–æ–¥–∏—Ç—Å—è –Ω–∞ –∑–∞–¥–Ω–µ–º –∫–æ–Ω—Ü–µ —Ç–µ–ª–∞. –ü–∞—Ä–∞–∑–∏—Ç–∏—Ä—É—é—Ç –≤ –∫–∏—à–µ—á–Ω–∏–∫–µ —Ä–µ–ø—Ç–∏–ª–∏–π, –ø—Ç–∏—Ü –∏ –º–ª–µ–∫–æ–ø–∏—Ç–∞—é—â–∏—Ö. –ü–µ—Ä–≤—ã–º–∏ –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω—ã–º–∏ —Ö–æ–∑—è–µ–≤–∞–º–∏ —è–≤–ª—è—é—Ç—Å—è –±—Ä—é—Ö–æ–Ω–æ–≥–∏–µ –º–æ–ª–ª—é—Å–∫–∏, –≤—Ç–æ—Ä—ã–º–∏ ‚Äì –±–µ—Å–ø–æ–∑–≤–æ–Ω–æ—á–Ω—ã–µ (–º–æ–ª–ª—é—Å–∫–∏, –ø–∏—è–≤–∫–∏, –æ–ª–∏–≥–æ—Ö–µ—Ç—ã) –∏ –ø–æ–∑–≤–æ–Ω–æ—á–Ω—ã–µ –≤—Å–µ—Ö –∫–ª–∞—Å—Å–æ–≤. –û—Ç—Ä—è–¥ –≤–∫–ª—é—á–∞–µ—Ç –¥–≤–∞ –ø–æ–¥–æ—Ç—Ä—è–¥–∞ Strigeata, Cyatocotylata, –∏–∑ –∫–æ—Ç–æ—Ä—ã—Ö –Ω–∞–∏–±–æ–ª—å—à–µ–µ –≤–µ—Ç–µ—Ä–∏–Ω–∞—Ä–Ω–æ–µ –∑–Ω–∞—á–µ–Ω–∏–µ –∏–º–µ–µ—Ç –ø–µ—Ä–≤—ã–π –ø–æ–¥–æ—Ç—Ä—è–¥, –≤–∫–ª—é—á–∞—é—â–∏–π –≤–æ—Å–µ–º—å —Å–µ–º–µ–π—Å—Ç–≤.
–°–µ–º. Strigeidae Railliet, 1919 ‚Äì —á–∞—à–µ–≤–∏–¥–Ω—ã–π –ø–µ—Ä–µ–¥–Ω–∏–π —Å–µ–≥–º–µ–Ω—Ç –æ—Ç–¥–µ–ª–µ–Ω –æ—Ç –∑–∞–¥–Ω–µ–≥–æ —Å—É–∂–µ–Ω–∏–µ–º –∏–ª–∏ –ø–µ—Ä–µ—Ç—è–∂–∫–æ–π. –ü–æ–ª–æ–≤–æ–µ –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ –≤–±–ª–∏–∑–∏ –∑–∞–¥–Ω–µ–≥–æ –∫–æ–Ω—Ü–∞ —Ç–µ–ª–∞. –°–µ–º–µ–Ω–Ω–∏–∫–∏ –ø–æ–∑–∞–¥–∏ —è–∏—á–Ω–∏–∫–∞, –º–∞—Ç–∫–∞ –∫–æ—Ä–æ—Ç–∫–∞—è (—Å–æ–¥–µ—Ä–∂–∏—Ç –º–∞–ª–æ —è–∏—Ü), –ø–µ—Ç–ª–∏ –µ–µ –ø—Ä–æ—Å—Ç–∏—Ä–∞—é—Ç—Å—è –Ω–µ –¥–∞–ª—å—à–µ –º–µ–∂—Å–µ–≥–º–µ–Ω—Ç–Ω–æ–π –≥—Ä–∞–Ω–∏—Ü—ã. –ú–∞—Ä–∏—Ç—ã –ø–∞—Ä–∞–∑–∏—Ç–∏—Ä—É—é—Ç –≤ –∫–∏—à–µ—á–Ω–∏–∫–µ –ø—Ç–∏—Ü. –Ý–∞–∑–≤–∏–≤–∞—é—Ç—Å—è —Å –¥–≤—É–º—è –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω—ã–º–∏ —Ö–æ–∑—è–µ–≤–∞–º–∏. –í—Ç–æ—Ä—ã–º –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω—ã–º —Ö–æ–∑—è–∏–Ω–æ–º —è–≤–ª—è—é—Ç—Å—è –º–æ–ª–ª—é—Å–∫–∏, –ø–∏—è–≤–∫–∏, —Ä—ã–±—ã, —Ä–µ–ø—Ç–∏–ª–∏–∏, –ø—Ç–∏—Ü—ã. –°–µ–º–µ–π—Å—Ç–≤–æ –æ–±—ä–µ–¥–∏–Ω—è–µ—Ç –æ–∫–æ–ª–æ 110 –≤–∏–¥–æ–≤, –Ω–∞–∏–±–æ–ª–µ–µ —Ä–∞—Å–ø—Ä–æ—Å—Ç—Ä–∞–Ω–µ–Ω—ã –≤–∏–¥—ã –∏–∑ —Ä–æ–¥–æ–≤ Strigea, Cotilurus, Apatemon.
Сем.Diplostomatidae (Poirier,1886) – рожковидные и лопатовидные трематоды, орган Брандеса в виде округлой присоски. На передней крае тела имеются псевдоприсоски. Матка и семяизвергательный канал, слмваются в непарный гермафродитный канал, открывающийся в полость полового атриума. Семейство включает 23 рода, наибольшее ветеринарное значение имеют представителей родов Diplostomum, Neodiplostomum, Postodiplostomum, Tylodelphue. Мариты паразитируют в кишечнике приводных птиц, первый промежуточный хозяин – пресноводные моллюски, второй – многие пресноводные рыбы, в хрусталике которых формируются метарцеркарии.
–°–µ–º. Alariidae-–º–µ–ª–∫–∏–µ —Ç—Ä–µ–º–∞—Ç–æ–¥—ã(2,4-4,4), –≤–æ–∫—Ä—É–≥ —Ä–æ—Ç–æ–≤–æ–π –ø—Ä–∏—Å–æ—Å–∫–∏ –∏–º–µ—é—Ç—Å—è —É—à–∫–æ–≤–∏–¥–Ω—ã–µ –≤—ã—Ä–æ—Å—Ç—ã. –ú–∞—Ä–∏—Ç—ã-Alaria alata-–ø–∞—Ä–∞–∑–∏—Ç–∏—Ä—É—é—Ç –≤ —Ç–æ–Ω–∫–æ–º –∫–∏—à–µ—á–Ω–∏–∫–µ –ø–ª–æ—Ç–æ—è–¥–Ω—ã—Ö. –Ý–∞–∑–≤–∏–≤–∞—é—Ç—Å—è —Å —É—á–∞—Å—Ç–∏–µ–º –¥–≤—É—Ö –ø—Ä–æ–º–µ–∂—É—Ç–æ—á–Ω—ã—Ö —Ö–æ–∑—è–µ–≤(–º–æ–ª–ª—é—Å–∫–∏, –∞–º—Ñ–∏–±–∏–∏).
–§–ê–°–¶–ò–û–õ–Å–ó–´ –ñ–ò–í–û–¢–ù–´–•
Фасциолёзы – инвазионные болезни домашних и диких животных, а также человека, вызываемые паразитированием в желчных ходах печени и желчном пузыре трематод Fasciola hepatica, F. gigantia из рода Fasciola, сем. Fasciolidae, подотряда Fasciolata, отряда Fasciolida. Fasciola hepatica – листовидная трематода, 20-30 мм длины и 8-12 мм ширины. Тегумент покрыт мелкими шипиками. В передней части располагаются ротовая и брюшная присоски, между ними открываются половые отверстия. Есть половая бурса. Позади брюшной присоски – матка, заполненная яйцами; слева от матки находится яичник. Среднюю и заднюю части тела паразита занимают два древовидно разветвленных семенника. Пищеварительная система начинается ротовым отверстием на дне ротовой присоски, затем следует глотка, короткий пищевод, переходящий в два кишечных ствола с боковыми ответвлениями. Fasciola gigantica – отличается от предыдущего вида большими размерами тела (длина 40-75 мм, ширина – 5-12 мм).
Биология возбудителей. Мариты откладывают в просвет желчных ходов яйца, которые затем с калом животных попадают во внешнюю среду. В воде при температуре 10-30 С0 и в темноте в яйце формируется мирацидий (0,15 мм длины), который покидает яйцо и в воде активно проникает в полость тела моллюсков. Промежуточными хозяевами F. hepatica являются малые прудовики - Lymnea truncatula, F. gigantica – ушковидные прудовики L. auricularia, где за 2-3 месяца проходят стадии развития (материнская спороциста, редия, церкария). Церкарии покидают моллюска, в воде некоторое время они свободно плавают, затем прикрепляются к какому-либо твердому предмету или остаются на поверхности воды и инцистируются, превращаясь в адолескариев.
–ñ–∏–≤–æ—Ç–Ω—ã–µ –∑–∞—Ä–∞–∂–∞—é—Ç—Å—è, –∑–∞–≥–ª–∞—Ç—ã–≤–∞—è —Å –≤–æ–¥–æ–π –∏–ª–∏ —Ç—Ä–∞–≤–æ–π —Ü–µ—Ä–∫–∞—Ä–∏–µ–≤ –∏–ª–∏ –∞–¥–æ–ª–µ—Å–∫–∞—Ä–∏–µ–≤. –í –æ—Ä–≥–∞–Ω–∏–∑–º–µ —Ö–æ–∑—è–µ–≤ –∞–¥–æ–ª–µ—Å–∫–∞—Ä–∏–∏ —Å–±—Ä–∞—Å—ã–≤–∞—é—Ç –æ–±–æ–ª–æ—á–∫—É, –ø—Ä–æ–Ω–∏–∫–∞—é—Ç –≤ –ø–∞—Ä–µ–Ω—Ö–∏–º—É –ø–µ—á–µ–Ω–∏ –≥–µ–º–∞—Ç–æ–≥–µ–Ω–Ω—ã–º –ø—É—Ç–µ–º –∏–ª–∏ —á–µ—Ä–µ–∑ —Å—Ç–µ–Ω–∫—É –∫–∏—à–µ—á–Ω–∏–∫–∞, —Ä–∞–∑—Ä—É—à–∞—é—Ç –ø–µ—á–µ–Ω–æ—á–Ω—É—é –∫–∞–ø—Å—É–ª—É. –í —Ç–µ—á–µ–Ω–∏–µ 35-40 –¥–Ω–µ–π –º–æ–ª–æ–¥—ã–µ —Ñ–∞—Å—Ü–∏–æ–ª—ã –º–∞–ª–æ–ø–æ–¥–≤–∏–∂–Ω—ã –∏ –≤—ã–∑—ã–≤–∞—é—Ç –Ω–µ–∑–Ω–∞—á–∏—Ç–µ–ª—å–Ω—ã–µ –ø–æ–≤—Ä–µ–∂–¥–µ–Ω–∏—è –ø–∞—Ä–µ–Ω—Ö–∏–º—ã. –° 38 –ø–æ 60-–π –¥–µ–Ω—å –ø—Ä–æ–∏—Å—Ö–æ–¥–∏—Ç –∞–∫—Ç–∏–≤–Ω–∞—è –º–∏–≥—Ä–∞—Ü–∏—è –Ω–µ–ø–æ–ª–æ–≤–æ–∑—Ä–µ–ª—ã—Ö —Ñ–∞—Å—Ü–∏–æ–ª –≤ –∂–µ–ª—á–Ω—ã–µ –ø—Ä–æ—Ç–æ–∫–∏ –ø–µ—á–µ–Ω–∏, –≤—ã–∑—ã–≤–∞—è –æ—Å—Ç—Ä—É—é —Ñ–æ—Ä–º—É —Ç–µ—á–µ–Ω–∏—è —Ñ–∞—Å—Ü–∏–æ–ª–µ–∑–∞. –ß–µ—Ä–µ–∑ 2, 5-4 –º–µ—Å—è—Ü–∞ —Ñ–∞—Å—Ü–∏–æ–ª—ã –¥–æ—Å—Ç–∏–≥–∞—é—Ç –ø–æ–ª–æ–≤–æ–π –∑—Ä–µ–ª–æ—Å—Ç–∏. –ü–∞—Ä–∞–∑–∏—Ç–∏—Ä—É—é—Ç –≤ –∂–µ–ª—á–Ω—ã—Ö —Ö–æ–¥–∞—Ö 1-3 –≥–æ–¥–∞, –≤—ã–∑—ã–≤–∞—è —Ö—Ä–æ–Ω–∏—á–µ—Å–∫—É—é —Ñ–æ—Ä–º—É —Ç–µ—á–µ–Ω–∏—è —Ñ–∞—Å—Ü–∏–æ–ª–µ–∑–∞.
–ú–∞—Ç–µ—Ä–∏–∞–ª —Å —Å–∞–π—Ç–∞: http://www.vetlib.ru/parasitologie/page,1,227-parazitologiya-klass-trematod-metodicheskie.html
–°—Ç—Ä–æ–µ–Ω–∏–µ –ª–∏—á–∏–Ω–æ–∫ –∏ –ø–∞—Ä—Ç–µ–Ω–æ–≥–µ–Ω–µ—Ç–∏—á–µ—Å–∫–∏—Ö –ø–æ–∫–æ–ª–µ–Ω–∏–π —Ç—Ä–µ–º–∞—Ç–æ–¥:
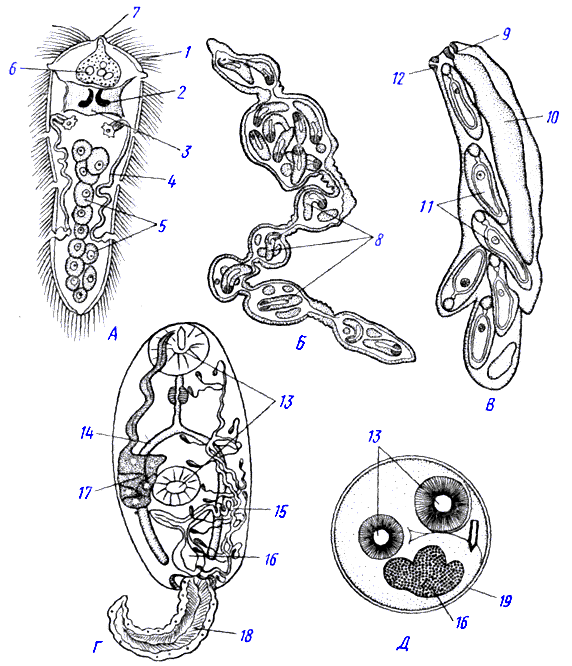
–°—Ç—Ä–æ–µ–Ω–∏–µ –ª–∏—á–∏–Ω–æ–∫ –∏ –ø–∞—Ä—Ç–µ–Ω–æ–≥–µ–Ω–µ—Ç–∏—á–µ—Å–∫–∏—Ö –ø–æ–∫–æ–ª–µ–Ω–∏–π —Ç—Ä–µ–º–∞—Ç–æ–¥ (–ø–æ —Ä–∞–∑–Ω—ã–º –∞–≤—Ç–æ—Ä–∞–º). –ê - –º–∏—Ä–∞—Ü–∏–¥–∏–π; –ë - —Å–ø–æ—Ä–æ—Ü–∏—Å—Ç–∞; –í - —Ä–µ–¥–∏—è; –ì -—Ü–µ—Ä–∫–∞—Ä–∏—è; –î - –º–µ—Ç–∞—Ü–µ—Ä–∫–∞—Ä–∏—è: 1 - —Ä–µ—Å–Ω–∏—á–∫–∏, 2 - –≥–ª–∞–∑–∞, 3 - –º–æ–∑–≥–æ–≤–æ–π –≥–∞–Ω–≥–ª–∏–π, 4 - –ø—Ä–æ—Ç–æ–Ω–µ—Ñ—Ä–∏–¥–∏–∏, 5 - –∑–∞—Ä–æ–¥—ã—à–µ–≤—ã–µ –∫–ª–µ—Ç–∫–∏, 6 - –∂–µ–ª–µ–∑–∞ –º–∏—Ä–∞—Ü–∏–¥–∏—è, 7 - —Ö–æ–±–æ—Ç–æ–∫, 8 - –∑–∞—Ä–æ–¥—ã—à–∏ —Ä–µ–¥–∏–π, 9 - –≥–ª–æ—Ç–∫–∞, 10 - –º–µ—à–∫–æ–≤–∏–¥–Ω—ã–π –∫–∏—à–µ—á–Ω–∏–∫, 11 - –∑–∞—Ä–æ–¥—ã—à–∏ —Ü–µ—Ä–∫–∞—Ä–∏–π, 12 - –æ—Ç–≤–µ—Ä—Å—Ç–∏–µ –¥–ª—è –≤—ã—Ö–æ–¥–∞ –∑—Ä–µ–ª—ã—Ö —Ü–µ—Ä–∫–∞—Ä–∏–π, 13 - —Ä–æ—Ç–æ–≤–∞—è –∏ –±—Ä—é—à–Ω–∞—è –ø—Ä–∏—Å–æ—Å–∫–∏, 14 - –∫–∏—à–µ—á–Ω–∏–∫, 15 - –≤—ã–¥–µ–ª–∏—Ç–µ–ª—å–Ω—ã–µ –∫–∞–Ω–∞–ª—ã, 16 - –º–æ—á–µ–≤–æ–π (—ç–∫—Å–∫—Ä–µ—Ç–æ—Ä–Ω—ã–π) –ø—É–∑—ã—Ä—å, 17 - –∂–µ–ª–µ–∑—ã –ø—Ä–æ–Ω–∏–∫–Ω–æ–≤–µ–Ω–∏—è, 18 - —Ö–≤–æ—Å—Ç —Ü–µ—Ä–∫–∞—Ä–∏–π, 19 - –æ–±–æ–ª–æ—á–∫–∞ —Ü–∏—Å—Ç—ã –º–µ—Ç–∞—Ü–µ—Ä–∫–∞—Ä–∏–∏

–¶–µ—Ä–∫–∞—Ä–∏—è —Ç—Ä–µ–º–∞—Ç–æ–¥—ã Fascioloides magna

–Ý–∏—Å. 15. –Ý–∞–∑–ª–∏—á–Ω—ã–µ —Ç–∏–ø—ã —Ü–µ—Ä–∫–∞—Ä–∏–µ–≤ —Ç—Ä–µ–º–∞—Ç–æ–¥ : 1 - –æ–¥–Ω–æ–ø—Ä–∏—Å–æ—Å–∫–æ–≤—ã–µ, 2 -
–∑–∞–¥–Ω–µ–ø—Ä–∏—Å–æ—Å–∫–æ–≤—ã–µ, 3 - –ø–ª–∞–≤–Ω–∏–∫–æ–≤–æ-–≥—Ä–µ–±–µ–Ω—á–∞—Ç—ã–µ (Opisthorchis felineus), 4 -
–Ω–µ–≤–æ–æ—Ä—É–∂–µ–Ω–Ω—ã–µ (Fasciola hepatica), 5 - –±—É–ª–∞–≤–æ—Ö–≤–æ—Å—Ç—ã–µ, 6 - —ç—Ö–∏–Ω–æ—Å—Ç–æ–º–∞—Ç–∏–¥–Ω—ã–µ, 7
- –º–µ—à–∫–æ—Ö–≤–æ—Å—Ç—ã–µ, 8 - –∫–æ—Ç–∏–ª—é—Ä–Ω—ã–µ, 9 - –≤–∏–ª–æ—Ö–≤–æ—Å—Ç—ã–µ, 10 - –∫–æ—Ä–æ—Ç–∫–æ—Ö–≤–æ—Å—Ç—ã–µ, 11 -
–±–µ—Å—Ö–≤–æ—Å—Ç—ã–µ, 12 - —Å—Ç–∏–ª–µ—Ç–Ω—ã–µ, 13 - —â–µ—Ç–∏–Ω–∫–æ—Ö–≤–æ—Å—Ç—ã–µ (–ø–æ –®—É–ª—å—Ü—É , –ì–≤–æ–∑–¥–µ–≤—É, 1970).
–° —Å–∞–π—Ç–∞: http://bio.ssmu.ru/faculty/Phis/materials/sosal.htm
–°—Å—ã–ª–∫–∏:
–°—Å—ã–ª–∫–∏:
1. –¶–µ—Ä–∫–∞—Ä–∏–∏
2. –ö–ª–∞—Å—Å —Å–æ—Å–∞–ª—å—â–∏–∫–∏: –∂–∏–∑–Ω–µ–Ω—ã–π —Ü–∏–∫–ª: –æ–±—â–∏–µ —Å–≤–µ–¥–µ–Ω–∏—è
3. –°–ø–æ—Ä–æ—Ü–∏—Å—Ç–∞
4. –Ý–µ–¥–∏—è
5. –ú–∏—Ä–∞—Ü–∏–¥–∏–π
#1 by Richard Mussell - –ú–∞—Ä—Ç 18th, 2013 at 15:00
What a superb write up