1.2.1. –í–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–∞—è –∏ –ø–æ–ª–æ–≤–∞—è —Ñ–∞–∑—ã
–≤ –∂–∏–∑–Ω–µ–Ω–Ω–æ–º —Ü–∏–∫–ª–µ
–º–Ω–æ–≥–æ–∫–ª–µ—Ç–æ—á–Ω—ã—Ö –æ—Ä–≥–∞–Ω–∏–∑–º–æ–≤
Vegetative and sexual phase in the life cycle of multicellular organisms
В клеточных популяциях прокариотов - бактерий и синезелёных водорослей - свойство неограниченной пролиферации присуще всем клеткам. Это же свойство наблюдается у простейших, которые относятся к эукариотам. Прокариоты и простейшие проходят и жизненном цикле несколько фаз, с которыми связано изменение их морфологических и поведенческих свойств. Некоторые простейшие (например, отдельные разновидности Sporozoa) в разных фазах образуют периодически то агрегированную многоклеточную, то одноклеточную форму [31]. В одну из фаз, связанную с дезагрегацией на отдельные клетки, образуются не споры, а половые клетки. У прокариотов и Sporozoa не образуются субпопуляции дифференцированных и детерминированных соматических клеток с ограниченной пролиферацией, как у более высокоорганизованных многоклеточных. Дифференцировка у них происходит циклически: в разных фазах своих жизненных циклов, а также под влиянием различных модифицирующих воздействий внешней среды в клетках развиваются специальные структуры, характеризующие то или иное дифференцированное состояние (например, флагеллы, псевдоподии, оболочки цист, реснички, раковины и т. д.). Все эти структуры непостоянны и при смене фаз жизненно­го цикла либо при изменениях внешних условий быстро утрачиваются и сменяются другими. Важная особенность прокариотов и простейших та, что все гены их геномов участвуют в реализации жизненного цикла [32].
В многоклеточных организмах ситуация более сложная. В основе их размножения лежит особая иммортализованная популяция клеток, существование которой было открыто в последнем десятилетии прошлого века |33|. В 1893 г. August Weisman [34] опубликовал «теорию зародышевой плазмы». Он обратил внимание на то, что у зародышей животных на разных стадиях развития обособляется группа клеток, которые у взрослого организма дают начало репродуктивным тканям. Эти клетки были названы автором «непрерывной линией половых клеток». В настоящее время существование непрерывной линии половых клеток подтверждено большим числом экспериментальных данных [35-37]. Непрерывная линия половых клеток - это реальный факт природы, и потому вполне обосновано и противопоставление друг другу сомы и этой бессмертной клеточной линии [6].
|
|
|
|
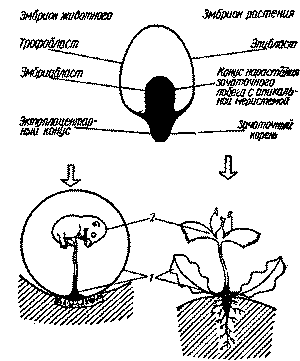
В жизненном цикле всех многоклеточных эукариотов (включая, по нашему мнению, и плацентарных млекопитающих) происходит смена исконных фаз жизненного цикла - вегетативной и половой. Вегетативная фаза может завершаться образованием новых - вегетативных - и половых организмов (при переходе в половую фазу). В половой фазе происходит половое размножение, причем из зиготы всегда образуется вегетативный организм. Эти особенности жизненных циклов присущи в равной мере многоклеточным животным и растениям (рис. 1). В составе тканей всех многоклеточных растений есть стойко воспроизводящая себя субпопуляция специальных клеток, которая по мере нормальной пролиферации «выделяет» из своего состава часть клеток, проходящих соматическую дифференцировку, образуя ткани и органы растения. Эту субпопуляцию клеток в тканях высших растений называют меристемой [38]. В верхушечной точке роста побега растения функционирует
–∞–ø–∏–∫–∞–ª—å–Ω–∞—è –º–µ—Ä–∏—Å—Ç–µ–º–∞, –∫–æ—Ç–æ—Ä–∞—è –≤ —Ä–µ–∑—É–ª—å—Ç–∞—Ç–µ —Å–≤–æ–µ–π –≤—ã—Å–æ–∫–æ—É–ø–æ—Ä—è–¥–æ—á–µ–Ω–Ω–æ–π –ø—Ä–æ–ª–∏—Ñ–µ—Ä–∞—Ü–∏–∏ –æ–±—Ä–∞–∑—É–µ—Ç –≤—Å–µ —Å–æ–º–∞—Ç–∏—á–µ—Å–∫–∏–µ –∫–ª–µ—Ç–æ—á–Ω—ã–µ —Ç–∏–ø—ã —Ç–∫–∞–Ω–µ–π —Ä–∞—Å—Ç–µ–Ω–∏—è, –∞ —Ç–∞–∫–∂–µ —Å—Ç–æ–π–∫–æ –∏ –Ω–µ–æ–≥—Ä–∞–Ω–∏—á–µ–Ω–Ω–æ –≤–æ—Å–ø—Ä–æ–∏–∑–≤–æ–¥–∏—Ç —Å–≤–æ—é —Å–æ–±—Å—Ç–≤–µ–Ω–Ω—É—é —Å—É–±–ø–æ–ø—É–ª—è—Ü–∏—é. –ê–ø–∏–∫–∞–ª—å–Ω–æ–π –º–µ—Ä–∏—Å—Ç–µ–º–µ —Å–≤–æ–π—Å—Ç–≤–µ–Ω–Ω–æ –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–µ —Ä–∞–∑–º–Ω–æ–∂–µ–Ω–∏–µ: –ø—Ä–∏ –æ—Ç—Ä–∞—Å—Ç–∞–Ω–∏–∏ –ø–æ–±–µ–≥–∞ –æ–Ω–∞ –æ—Å—Ç–∞–≤–ª—è–µ—Ç –≤ –µ–≥–æ –æ—Ç–¥–µ–ª—å–Ω—ã—Ö —É—á–∞—Å—Ç–∫–∞—Ö –∑–∞–ø–∞—Å–Ω—ã–µ –º–µ—Ä–∏—Å—Ç–µ–º—ã - –ø–æ—á–∫–∏, –∞ —Ç–∞–∫–∂–µ –∫–∞–º–±–∏–∞–ª—å–Ω—ã–µ –∏ –¥—Ä—É–≥–∏–µ –∫–ª–µ—Ç–∫–∏ —Å–æ–º—ã, –∫–æ—Ç–æ—Ä—ã–µ –ø—Ä–∏ –æ–ø—Ä–µ–¥–µ–ª–µ–Ω–Ω—ã—Ö —ç–∫—Å—Ç—Ä–µ–º–∞–ª—å–Ω—ã—Ö —É—Å–ª–æ–≤–∏—è—Ö –º–æ–≥—É—Ç –æ–±—Ä–∞–∑–æ–≤—ã–≤–∞—Ç—å –∫–∞–ª–ª—é—Å—ã –∏ –Ω–æ–≤—ã–µ –º–µ—Ä–∏—Å—Ç–µ–º–∞–ª—å–Ω—ã–µ –∑–æ–Ω—ã [38|. –ü—Ä–∏ —Å–º–µ–Ω–µ –æ—á–µ—Ä–µ–¥–Ω–æ–π —Ñ–∞–∑—ã –∂–∏–∑–Ω–µ–Ω–Ω–æ–≥–æ —Ü–∏–∫–ª–∞ –º–µ—Ä–∏—Å—Ç–µ–º–∞ –¥–µ–∑–∞–≥—Ä–µ–≥–∏—Ä—É–µ—Ç—Å—è –Ω–∞ –æ—Ç–¥–µ–ª—å–Ω—ã–µ –∫–ª–µ—Ç–∫–∏ - —Å–ø–æ—Ä—ã, –∫–æ—Ç–æ—Ä—ã–µ –≤ —Ä–µ–∑—É–ª—å—Ç–∞—Ç–µ —Ä—è–¥–∞ –ø—Ä–µ–æ–±—Ä–∞–∑–æ–≤–∞—Ç–µ–ª—å–Ω—ã—Ö –ø—Ä–æ—Ü–µ—Å—Å–æ–≤ (—Ä–∞–∑–ª–∏—á–Ω—ã—Ö —É —Ä–∞–∑–Ω—ã—Ö —ç–≤–æ–ª—é—Ü–∏–æ–Ω–Ω—ã—Ö –≥—Ä—É–ø–ø —Ä–∞—Å—Ç–µ–Ω–∏–π) –ø–µ—Ä–µ—Ö–æ–¥—è—Ç –≤ –ø–æ–ª–æ–≤—É—é —Ñ–∞–∑—É - –æ–±—Ä–∞–∑—É—é—Ç –ø–æ–ª–æ–≤—ã–µ –∫–ª–µ—Ç–∫–∏ –∏ –≥–æ–Ω–∞–¥—ã. –ø–æ–ª–æ–≤—ã–µ –∫–ª–µ—Ç–∫–∏ —Å–ª–∏–≤–∞—é—Ç—Å—è –≤ –∑–∏–≥–æ—Ç—É, –∫–æ—Ç–æ—Ä–∞—è –ø—Ä–µ–¥—Å—Ç–∞–≤–ª—è–µ—Ç —Å–æ–±–æ–π –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω—ã–π –æ—Ä–≥–∞–Ω–∏–∑–º –∏ –¥–∞–µ—Ç –Ω–∞—á–∞–ª–æ –∞–≥—Ä–µ–≥–∏—Ä–æ–≤–∞–Ω–Ω–æ–π –∞–ø–∏–∫–∞–ª—å–Ω–æ–π –º–µ—Ä–∏—Å—Ç–µ–º–µ.
–°—Ö–æ–¥–Ω—ã–º –æ–±—Ä–∞–∑–æ–º –ø—Ä–æ–∏—Å—Ö–æ–¥–∏—Ç —Å–º–µ–Ω–∞ –æ—Å–Ω–æ–≤–Ω—ã—Ö —Ñ–∞–∑ –∏ –≤ –∂–∏–∑–Ω–µ–Ω–Ω–æ–º —Ü–∏–∫–ª–µ –∂–∏–≤–æ—Ç–Ω—ã—Ö. –°—Ä–∞–≤–Ω–∏–≤–∞—è —Ä–∞–∑–ª–∏—á–Ω—ã–µ –≤–∏–¥—ã (–º–Ω–æ–≥–æ–∫–ª–µ—Ç–æ—á–Ω—ã—Ö) –æ—Ä–≥–∞–Ω–∏–∑–º–æ–≤ –≤ —Å–æ–æ—Ç–≤–µ—Ç—Å—Ç–≤—É—é—â–∏—Ö –æ—Å–Ω–æ–≤–Ω—ã—Ö —Ñ–∞–∑–∞—Ö, –º—ã –æ–±–Ω–∞—Ä—É–∂–∏–≤–∞–µ–º —Ä—è–¥ –æ–±—â–∏—Ö –º–æ—Ä—Ñ–æ–ª–æ–≥–∏—á–µ—Å–∫–∏—Ö —á–µ—Ä—Ç: –∏–∑ –∑–∏–≥–æ—Ç—ã –æ–±—Ä–∞–∑—É–µ—Ç—Å—è –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω—ã–π –æ—Ä–≥–∞–Ω–∏–∑–º, –ø—Ä–∏—á–µ–º –º–æ—Ä—Ñ–æ–ª–æ–≥–∏—á–µ—Å–∫–∏ —ç—Ç–∏ –≤–æ–∑–Ω–∏–∫–∞—é—â–∏–µ –∏–∑ –∑–∏–≥–æ—Ç—ã –æ—Ä–≥–∞–Ω–∏–∑–º—ã –≤–æ –º–Ω–æ–≥–æ–º —Å—Ö–æ–¥–Ω—ã —É —Ä–∞—Å—Ç–µ–Ω–∏–π –∏ –∂–∏–≤–æ—Ç–Ω—ã—Ö. –ú–µ—Ö–∞–Ω–∏–∑–º –∏–º–ø–ª–∞–Ω—Ç–∞—Ü–∏–∏ –±–ª–∞—Å—Ç–æ—Ü–∏—Å—Ç—ã –∂–∏–≤–æ—Ç–Ω—ã—Ö (–æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏–µ —ç–∫—Ç–æ–ø–ª–∞—Ü–µ–Ω—Ç–∞—Ä–Ω–æ–≥–æ –∫–æ–Ω—É—Å–∞) –Ω–∞–ø–æ–º–∏–Ω–∞–µ—Ç –º–µ—Ö–∞–Ω–∏–∑–º —É–∫–æ—Ä–µ–Ω–µ–Ω–∏—è —Å–µ–º–µ–Ω–∏ —Ä–∞—Å—Ç–µ–Ω–∏–π (–æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏–µ –∑–∞—á–∞—Ç–æ—á–Ω–æ–≥–æ –∫–æ—Ä–Ω—è) (—Å–º. —Ä–∏—Å. 1 –∏ 2). –û—Å—Ç–∞—ë—Ç—Å—è –æ—Ç–∫—Ä—ã—Ç—ã–º –≤–æ–ø—Ä–æ—Å, —è–≤–ª—è–µ—Ç—Å—è –ª–∏ —ç—Ç–æ —Å—Ö–æ–¥—Å—Ç–≤–æ –æ—Ä–≥–∞–Ω–∏–∑–º–æ–≤
|
|
|
–Ý–∏—Å. 2, –ë–ª–∞—Å—Ç–æ—Ü–∏—Å—Ç–∞ –∫—Ä—ã—Å—ã: a - –Ω–∞—á–∞–ª–æ –∏–º–ø–ª–∞–Ω—Ç–∞—Ü–∏–∏ –± - —Å—Ç–∞–¥–∏—è –¥–≤—É—Ö—Å–ª–æ–π–Ω–æ–≥–æ —è–π—Ü–µ–≤–æ–≥–æ —Ü–∏–ª–∏–Ω–¥—Ä–∞; 1 - –Ω–º–ø–ª–∞–Ω—Ç–∞—Ü–∏–æ–Ω–Ω–∞—è –∫—Ä–∏–ø—Ç–∞; 2 - –∑–∫—Ç–æ–ø–ª–∞—Ü–µ–Ω—Ç–∞—Ä–Ω—ã–π –∫–æ–Ω—É—Å; 3 - –∑–∞—Ä–æ–¥—ã—à–µ–≤–∞—è —ç–∫—Ç–æ–¥–µ—Ä–º–∞, 4 - –ø—Ä–æ–∫—Å–∏–º–∞–ª—å–Ω–∞—è –µ–Ω—Ç–æ–¥–µ—Ä–º–∞; 5 - –¥–∏—Å—Ç–∞–ª—å–Ω–∞—è –µ–Ω—Ç–æ–¥–µ—Ä–º–∞; 6 - —Ç—Ä–æ—Ñ–æ–±–ª–∞—Å—Ç; 7 - ¬´–∂–µ–ª—Ç–æ—á–Ω–∞—謪 –ø–æ–ª–æ—Å—Ç—å; 8 -–ø—Ä–æ–∫—Å–∏–º–∞–ª—å–Ω–∞—è –µ–Ω—Ç–æ–¥–µ—Ä–º–∞; 9 -–≤–Ω–µ–∑–∞—Ä–æ–¥—ã—à–µ–≤–∞—è (–¥–∏—Å—Ç–∞–ª—å–Ω–∞—è) —ç–∫—Ç–æ–¥–µ—Ä–º–∞; 10 - –∫–ª–µ—Ç–∫–∏ –¥–∏—Å—Ç–∞–ª—å–Ω–æ–π —ç–∫—Ç–æ–¥–µ—Ä–º—ã; 11 - –ø—Ä–æ–∞–º–Ω–∏–æ—Ç–∏—á–µ—Å–∫–∞—è –ø–æ–ª–æ—Å—Ç—å 12 - —Ç—Ä–æ—Ñ–æ—ç–∫—Ç–æ–¥–µ—Ä–º–∞ |


–≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω—ã—Ö —Ñ–∞–∑ –∞–Ω–∞–ª–æ–≥–∏–µ–π –∏–ª–∏ –≥–æ–º–æ–ª–æ–≥–∏–µ–π. –í –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π —Ñ–∞–∑–µ –æ—Ä–≥–∞–Ω–∏–∑–º, —Ä–∞–∑–≤–∏–≤–∞—è—Å—å, –º–æ–∂–µ—Ç –ø—Ä–æ–π—Ç–∏ –æ—Ç –æ–¥–Ω–æ–≥–æ –¥–æ –Ω–µ—Å–∫–æ–ª—å–∫–∏—Ö (–∞ –ø—Ä–∏ —Å–æ–æ—Ç–≤–µ—Ç—Å—Ç–≤—É—é—â–∏—Ö —É—Å–ª–æ–≤–∏—è—Ö –Ω–µ–æ–≥—Ä–∞–Ω–∏—á–µ–Ω–Ω–æ–µ –∫–æ–ª–∏—á–µ—Å—Ç–≤–æ) —Ü–∏–∫–ª–æ–≤ –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–≥–æ —Ä–∞–∑–º–Ω–æ–∂–µ–Ω–∏—è, –∏ –∑–∞—Ç–µ–º –æ–±—Ä–∞–∑—É–µ—Ç –æ—Ä–≥–∞–Ω–∏–∑–º –≤ –ø–æ–ª–æ–≤–æ–π —Ñ–∞–∑–µ. –ü—Ä–∏ —ç—Ç–æ–º –æ–±—Ä–∞–∑—É—é—Ç—Å—è –≥–æ–Ω–∞–¥—ã, –≤ –∫–æ—Ç–æ—Ä—ã–µ –≤—ã—Å–µ–ª—è—é—Ç—Å—è –ø—Ä–µ–¥—à–µ—Å—Ç–≤–µ–Ω–Ω–∏–∫–∏ –ø–æ–ª–æ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫.
|
|
|
–Ý–∏—Å. 3. –ù–µ–ø—Ä–µ—Ä—ã–≤–Ω–∞—è –ª–∏–Ω–∏—è –ø–æ–ª–æ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫ –≤ –∂–∏–∑–Ω–µ–Ω–Ω–æ–º —Ü–∏–∫–ª–µ –ø–ª–∞—Ü–µ–Ω—Ç–∞—Ä–Ω—ã—Ö –º–ª–µ–∫–æ–ø–∏—Ç–∞—é—â–∏—Ö: 1 -–≤—ã—Å–µ–ª–µ–Ω–∏–µ –≥–æ–Ω–æ—Ü–∏—Ç–æ–≤ –∏–∑ –∂–µ–ª—Ç–æ—á–Ω–æ–≥–æ –º–µ—à–∫–∞; 2 - –º–µ–π–æ–∑; –ó - —Å–ª–∏—è–Ω–∏–µ –ø–æ–ª–æ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫ —Å –æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏–µ–º –∑–∏–≥–æ—Ç—ã; 4 - –Ω–∞—á–∞–ª–æ –¥—Ä–æ–±–ª–µ–Ω–∏—è –∑–∏–≥–æ—Ç—ã –∏ –æ–±–æ—Å–æ–±–ª–µ–Ω–∏—è —Å–æ–º—ã –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π —Ñ–∞–∑—ã; 5 - –ø–µ—Ä–∏–æ–¥ —Ä–∞–∑–≤–∏—Ç–∏—è –æ—Ä–≥–∞–Ω–∏–∑–º–∞ –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π —Ñ–∞–∑—ã; 6-–ø–µ—Ä–∏–æ–¥ –∑–∞–∫–ª–∞–¥–∫–∏ —Å–æ–º—ã –æ—Ä–≥–∞–Ω–∏–∑–º–∞ –ø–æ–ª–æ–≤–æ–π —Ñ–∞–∑—ã; 7 - –ø–µ—Ä–∏–æ–¥ —Ñ—É–Ω–∫—Ü–∏–æ–Ω–∏—Ä–æ–≤–∞–Ω–∏—è ¬´–∞–ø–∏–∫–∞–ª—å–Ω–æ–π –º–µ—Ä–∏—Å—Ç–µ–º—㬪; 8 - —Ü–∏–∫–ª –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–≥–æ —Ä–∞–∑–º–Ω–æ–∂–µ–Ω–∏—è (–æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏–µ –æ–¥–Ω–æ—è–π—Ü–µ–≤—ã—Ö –±–ª–∏–∑–Ω–µ—Ü–æ–≤); 9 - —Ü–∏–∫–ª –ø–∞—Ä—Ç–µ–Ω–æ–≥–µ–Ω–µ—Ç–∏—á–µ—Å–∫–æ–≥–æ —Ä–∞–∑–º–Ω–æ–∂–µ–Ω–∏—è; 10 - –º–æ–º–µ–Ω—Ç —Ä–æ–¥–æ–≤; 11 - –ø–µ—Ä–∏–æ–¥ –ª–∞–∫—Ç–∞—Ü–∏–∏; 12 - –∑–∞–≤–µ—Ä—à–µ–Ω–∏–µ –ø–æ–ª–æ–≤–æ–≥–æ —Å–æ–∑—Ä–µ–≤–∞–Ω–∏—è –æ—Ä–≥–∞–Ω–∏–∑–º–∞ –ø–æ–ª–æ–≤–æ–π —Ñ–∞–∑—ã; 13 - —Å–º–µ—Ä—Ç—å –æ—Ä–≥–∞–Ω–∏–∑–º–∞ –ø–æ–ª–æ–≤–æ–π —Ñ–∞–∑—ã; 14 - —Å–æ–º–∞ –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π —Ñ–∞–∑—ã; 15 - —Å–æ–º–∞ –ø–æ–ª–æ–≤–æ–π —Ñ–∞–∑—ã; 15 - –ª–∏–Ω–∏—è –ø–æ–ª–æ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫ |

–ü—Ä–µ–¥—à–µ—Å—Ç–≤–µ–Ω–Ω–∏–∫–∏ –ø–æ–ª–æ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫ —É —Ä–∞—Å—Ç–µ–Ω–∏–π, –∫–∞–∫ —É–∂–µ —É–∫–∞–∑—ã–≤–∞–ª–æ—Å—å, –ø—Ä–æ–∏—Å—Ö–æ–¥—è—Ç –∏–∑ –∞–ø–∏–∫–∞–ª—å–Ω—ã—Ö –º–µ—Ä–∏—Å—Ç–µ–º –æ—Ä–≥–∞–Ω–∏–∑–º–∞ –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π —Ñ–∞–∑—ã, –∫–æ—Ç–æ—Ä—ã–µ –ø—Ä–∏ —ç—Ç–æ–º –¥–µ–∑–∞–≥—Ä–µ–≥–∏—Ä—É—é—Ç—Å—è. –£ –∂–∏–≤–æ—Ç–Ω—ã—Ö –≥–æ–Ω–æ—Ü–∏—Ç—ã –≤—ã—Å–µ–ª—è—é—Ç—Å—è –∏–∑ —Å–ø–µ—Ü–∏–∞–ª—å–Ω–æ–π –≥–µ—Ä–º–∏–Ω–∞—Ç–∏–≤–Ω–æ–π –∑–æ–Ω—ã –∂–µ–ª—Ç–æ—á–Ω–æ–≥–æ –º–µ—à–∫–∞ [35]. –ñ–µ–ª—Ç–æ—á–Ω—ã–π –º–µ—à–æ–∫ –º–ª–µ–∫–æ–ø–∏—Ç–∞—é—â–∏—Ö –º–æ–∂–Ω–æ —É–ø–æ–¥–æ–±–∏—Ç—å –ø–æ–±–µ–≥—É, –∞ —ç—Ç—É –æ—Å–æ–±—É—é –∑–æ–Ω—É —Å—á–∏—Ç–∞—Ç—å –∞–ø–∏–∫–∞–ª—å–Ω–æ–π –º–µ—Ä–∏—Å—Ç–µ–º–æ–π –∂–∏–≤–æ—Ç–Ω—ã—Ö.
–í —Å–æ—Å—Ç–∞–≤–µ —Ç–∫–∞–Ω–µ–π –æ—Ä–≥–∞–Ω–∏–∑–º–∞ –ø–æ–ª–æ–≤–æ–π —Ñ–∞–∑—ã –∞–ø–∏–∫–∞–ª—å–Ω–æ–π –º–µ—Ä–∏—Å—Ç–µ–º—ã –Ω–µ—Ç, –≤–æ—Å–ø—Ä–æ–∏–∑–≤–æ–¥—Å—Ç–≤–æ —Å–æ–º—ã –æ–±–µ—Å–ø–µ—á–∏–≤–∞–µ—Ç—Å—è –∫–∞–º–±–∏–∞–ª—å–Ω—ã–º–∏ –∫–ª–µ—Ç–∫–∞–º–∏, –≤—ã–¥–µ–ª—è—é—â–∏–º–∏ –∏–∑ —Å–≤–æ–µ–≥–æ —Å–æ—Å—Ç–∞–≤–∞ —Å—Ç–≤–æ–ª–æ–≤—ã–µ –∫–ª–µ—Ç–∫–∏ [39, 40]. –ì–µ—Ä–º–∏–Ω–∞—Ç–∏–≤–Ω—ã–µ –∫–ª–µ—Ç–∫–∏, –∑–∞—Å–µ–ª—è—é—â–∏–µ –≥–æ–Ω–∞–¥—ã, —Ñ–∞–∫—Ç–∏—á–µ—Å–∫–∏ –Ω–∞—Ö–æ–¥—è—Ç—Å—è –≤–Ω—É—Ç—Ä–∏ –ø–æ–ª–æ–≤—ã—Ö –ø—Ä–æ—Ç–æ–∫–æ–≤ –∏ –æ—Ç–≥—Ä–∞–Ω–∏—á–µ–Ω–∏–∏ –æ—Ç —Ç–∫–∞–Ω–µ–π –æ—Ä–≥–∞–Ω–∏–∑–º–∞ —ç–ø–∏—Ç–µ–ª–∏–∞–ª—å–Ω—ã–º –±–∞—Ä—å–µ—Ä–æ–º.
–¢–∞–∫–∏–º –æ–±—Ä–∞–∑–æ–º, –Ω–µ–ø—Ä–µ—Ä—ã–≤–Ω–∞—è –ª–∏–Ω–∏—è –ø–æ–ª–æ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫ –≤ —Å–≤–æ–µ–º —Ä–∞–∑–≤–∏—Ç–∏–∏ –ø—Ä–æ—Ö–æ–¥–∏—Ç —Å–æ–±—Å—Ç–≤–µ–Ω–Ω—ã–π –∑–∞–º–∫–Ω—É—Ç—ã–π –∂–∏–∑–Ω–µ–Ω–Ω—ã–π —Ü–∏–∫–ª (—Ä–∏—Å. 3), –ø–æ–¥–æ–±–Ω—ã–π –∂–∏–∑–Ω–µ–Ω–Ω—ã–º —Ü–∏–∫–ª–∞–º –ø—Ä–æ–∫–∞—Ä–∏–æ—Ç–æ–≤ –∏ Protozoa. –í —Ä–∞–∑–Ω—ã—Ö —Ñ–∞–∑–∞—Ö —ç—Ç–æ–≥–æ —Ü–∏–∫–ª–∞ –ø—Ä–æ–∏—Å—Ö–æ–¥—è—Ç –≤—Ä–µ–º–µ–Ω–Ω—ã–µ –¥–∏—Ñ—Ñ–µ—Ä–µ–Ω—Ü–∏—Ä–æ–≤–æ—á–Ω—ã–µ –∏–∑–º–µ–Ω–µ–Ω–∏—è –ø–æ–ª–æ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫ —Å –æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏–µ–º —Å–æ–æ—Ç–≤–µ—Ç—Å—Ç–≤—É—é—â–∏—Ö –≤—Ä–µ–º–µ–Ω–Ω—ã—Ö —Ñ—É–Ω–∫—Ü–∏–æ–Ω–∞–ª—å–Ω—ã—Ö —Å—Ç—Ä—É–∫—Ç—É—Ä - –ø—Å–µ–≤–¥–æ–ø–æ–¥–∏–π, —Ö–≤–æ—Å—Ç–æ–≤ —Å–ø–µ—Ä–º–∏–π, zona Pellucida –∏ —Ç. –¥. –ö –∏—Ö —á–∏—Å–ª—É, –ø–æ –Ω–∞—à–µ–º—É –º–Ω–µ–Ω–∏—é, —Å–ª–µ–¥—É–µ—Ç –æ—Ç–Ω–µ—Å—Ç–∏ –∏ —Å–æ–º—É, –¥–ª—è –æ–±—Ä–∞–∑–æ–≤–∞–Ω–∏—è –∫–æ—Ç–æ—Ä–æ–π –∂–µ—Ä—Ç–≤—É–µ—Ç—Å—è —á–∞—Å—Ç—å –∫–ª–µ—Ç–æ–∫ –Ω–µ–ø—Ä–µ—Ä—ã–≤–Ω–æ–π –ª–∏–Ω–∏–∏. –û—á–µ–≤–∏–¥–Ω–æ, –¥–ª—è –ø—Ä–æ—Ö–æ–∂–¥–µ–Ω–∏—è –Ω–µ–ø—Ä–µ—Ä—ã–≤–Ω–æ–≥–æ —Ü–∏–∫–ª–∞ –≤ –ª–∏–Ω–∏–∏ –ø–æ–ª–æ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫ –ø—Ä–æ–∏—Å—Ö–æ–¥–∏—Ç –ø–µ—Ä–∏–æ–¥–∏—á–µ—Å–∫–∞—è —ç–∫—Å–ø—Ä–µ—Å—Å–∏—è - —Ä–µ–ø—Ä–µ—Å—Å–∏—è –æ–ø—Ä–µ–¥–µ–ª–µ–Ω–Ω—ã—Ö –≥—Ä—É–ø–ø –≥–µ–Ω–æ–≤, –∫–æ—Ç–æ—Ä—ã–µ —Ñ—É–Ω–∫—Ü–∏–æ–Ω–∏—Ä—É—é—Ç —Ç–æ–ª—å–∫–æ –≤ –≥–µ—Ä–º–∏–Ω–∞—Ç–∏–≤–Ω—ã—Ö –∫–ª–µ—Ç–∫–∞—Ö. –î–ª—è –∫–ª–µ—Ç–æ–∫, –∫–æ—Ç–æ—Ä—ã–µ –≤—Å—Ç—É–ø–∞—é—Ç –Ω–∞ –ø—É—Ç—å —Å–æ–º–∞—Ç–∏—á–µ—Å–∫–æ–π –¥–µ—Ç–µ—Ä–º–∏–Ω–∞—Ü–∏–∏, –Ω–µ–æ–±—Ö–æ–¥–∏–º–∞ —ç–∫—Å–ø—Ä–µ—Å—Å–∏—è –µ—â–µ –∏ –º–Ω–æ–≥–∏—Ö –¥—Ä—É–≥–∏—Ö –≥—Ä—É–ø–ø –≥–µ–Ω–æ–≤, –∫–æ—Ç–æ—Ä—ã–µ –≤ –ª–∏–Ω–∏–∏ –ø–æ–ª–æ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫, –ø–æ-–≤–∏–¥–∏–º–æ–º—É, –Ω–∏–∫–æ–≥–¥–∞ –Ω–µ —ç–∫—Å–ø—Ä–µ—Å—Å–∏—Ä—É—é—Ç—Å—è. –ú–æ–∂–Ω–æ –ø—Ä–µ–¥–ø–æ–ª–æ–∂–∏—Ç—å, —á—Ç–æ –ø–æ—Å–ª–µ–¥–æ–≤–∞—Ç–µ–ª—å–Ω–æ—Å—Ç—å –≤–∫–ª—é—á–µ–Ω–∏—è –≥–µ–Ω–æ–≤ –≤ —Å–æ–º–∞—Ç–∏—á–µ—Å–∫–∏—Ö –∫–ª–µ—Ç–∫–∞—Ö –∏–º–µ–µ—Ç –Ω–µ —Ü–∏–∫–ª–∏—á–µ—Å–∫–∏–π —Ö–∞—Ä–∞–∫—Ç–µ—Ä, –∫–∞–∫ –≤ –Ω–µ–ø—Ä–µ—Ä—ã–≤¬≠–Ω–æ–π –ª–∏–Ω–∏–∏ –ø–æ–ª–æ–≤—ã—Ö –∫–ª–µ—Ç–æ–∫, –∞ –ª–∏–Ω–µ–π–Ω—ã–π: –æ—Ç –Ω–∞—á–∞–ª—å–Ω–æ–π, –∏–ª–∏ –∏–Ω–∏—Ü–∏–∞–ª—å–Ω–æ–π, –≥—Ä—É–ø–ø—ã –≥–µ–Ω–æ–≤ –∫ –∫–æ–Ω–µ—á–Ω–æ–π, –∏–ª–∏ —Ç–µ—Ä–º–∏–Ω–∞–ª—å–Ω–æ–π, –æ–±–µ—Å–ø–µ—á–∏–≤–∞—é—â–µ–π —Ñ—É–Ω–∫—Ü–∏—é ¬´—Ä–∞–±–æ—á–µ–𬪠—Å–º–µ—Ä—Ç–Ω–æ–π —Å–æ–º–∞—Ç–∏—á–µ—Å–∫–æ–π –∫–ª–µ—Ç–∫–∏. –≠—Ç–∞ —É—Å–ª–æ–≤–Ω–∞—è –¥–µ—Ç–µ—Ä–º–∏–Ω–∞—Ü–∏–æ–Ω–Ω–∞—è –ª–∏–Ω–∏—è –≥–µ–Ω–Ω–æ–π —ç–∫—Å–ø—Ä–µ—Å—Å–∏–∏ —Ä–∞–∑–≤–µ—Ç–≤–ª–µ–Ω–∞ –≤ —Å–æ–æ—Ç–≤–µ—Ç—Å—Ç–≤–∏–∏ —Å–æ “—Å—Ö–µ–º–æ–π” –¥–∏–≤–µ—Ä–≥–µ–Ω—Ü–∏–∏, –±–ª–∞–≥–æ–¥–∞—Ä—è —á–µ–º—É –æ–±—Ä–∞–∑—É—é—Ç—Å—è –¥–∏—Ñ—Ñ–µ—Ä–µ–Ω—Ü–∏—Ä–æ–≤–∞–Ω–Ω—ã–µ —Ñ—É–Ω–∫—Ü–∏–æ–Ω–∞–ª—å–Ω—ã–µ –∫–ª–µ—Ç–∫–∏ —Ä–∞–∑–Ω—ã—Ö —Ç–∏–ø–æ–≤ [40].
–ì–ª–∞–≤–Ω—ã–µ —Ñ–∞–∑—ã –∂–∏–∑–Ω–µ–Ω–Ω–æ–≥–æ —Ü–∏–∫–ª–∞ - –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–∞—è –∏ –ø–æ–ª–æ–≤–∞—è - —É —Ä–∞–∑–Ω—ã—Ö –∂–∏–≤–æ—Ç–Ω—ã—Ö –∏ —Ä–∞—Å—Ç–µ–Ω–∏–π –∏–º–µ—é—Ç —Ä–∞–∑–Ω—ã–π —É–¥–µ–ª—å–Ω—ã–π –≤–µ—Å, —á—Ç–æ –ø—Ä–æ—è–≤–ª—è–µ—Ç—Å—è –≤ —Å—Ç–µ–ø–µ–Ω–∏ —Ñ—É–Ω–∫—Ü–∏–æ–Ω–∞–ª—å–Ω–æ–π –¥–∏—Ñ—Ñ–µ—Ä–µ–Ω—Ü–∏—Ä–æ–≤–∫–∏ —Å–æ–º—ã –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–≥–æ –ª–∏–±–æ –ø–æ–ª–æ–≤–æ–≥–æ –æ—Ä–≥–∞–Ω–∏–∑–º–∞, –æ—Ç–Ω–æ—Å–∏—Ç–µ–ª—å–Ω–æ–π –¥–ª–∏—Ç–µ–ª—å–Ω–æ—Å—Ç–∏ –µ–≥–æ —Ñ—É–Ω–∫—Ü–∏–æ–Ω–∏—Ä–æ–≤–∞–Ω–∏—è [18]. –°—É—â–µ—Å—Ç–≤—É—é—Ç, –Ω–∞–ø—Ä–∏–º–µ—Ä, —Ç–∞–∫–∏–µ –≤–∏–¥—ã –º–µ—Ç–∞–≥–µ–Ω–µ—Ç–∏—á–µ—Å–∫–∏—Ö –∫–∏—à–µ—á–Ω–æ–ø–æ–ª–æ—Å—Ç–Ω—ã—Ö - –æ–¥–∏–Ω–æ—á–Ω—ã–µ –∏–ª–∏ –∫–æ–ª–æ–Ω–∏–∞–ª—å–Ω—ã–µ –ø–æ–ª–∏–ø—ã, –∂–∏–∑–Ω–µ–¥–µ—è—Ç–µ–ª—å–Ω–æ—Å—Ç—å –∫–æ—Ç–æ—Ä—ã—Ö –ø—Ä–æ—Ç–µ–∫–∞–µ—Ç –ø—Ä–µ–∏–º—É—â–µ—Å—Ç–≤–µ–Ω–Ω–æ –≤ –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π —Ñ–∞–∑–µ [42]. –ü–æ–ª–æ–≤—ã–µ –æ—Å–æ–±–∏ (–º–µ–¥—É–∑–æ–∏–¥—ã) —É –Ω–∏—Ö –æ–±—Ä–∞–∑—É—é—Ç—Å—è —Ä–µ–¥–∫–æ, —É –Ω–µ–∫–æ—Ç–æ—Ä—ã—Ö –∫–æ–ª–æ–Ω–∏–∞–ª—å–Ω—ã—Ö –≤–∏–¥–æ–≤ –≤–æ–æ–±—â–µ –Ω–µ –æ—Ç–¥–µ–ª—è—é—Ç—Å—è –æ—Ç –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π –∫–æ–ª–æ–Ω–∏–∏ (–ø–æ–¥–æ–±–Ω–æ —Ü–≤–µ—Ç–∞–º —É –≤—ã—Å—à–∏—Ö —Ä–∞—Å—Ç–µ–Ω–∏–π [12]. –î—Ä—É–≥–∞—è –∫—Ä–∞–π–Ω–æ—Å—Ç—å - —Å–Ω–∏–∂–µ–Ω–∏–µ —Ä–æ–ª–∏ –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π —Ñ–∞–∑—ã –∏ –ø—Ä–µ–≤–∞–ª–∏—Ä–æ–≤–∞–Ω–∏–µ –ø–æ–ª–æ–≤–æ–π. –¢–∞–∫, —Å—Ü–∏—Ñ–æ–∏–¥–Ω–∞—è –º–µ–¥—É–∑–∞ –ë—É–≥–µ–Ω–≤–∏–ª–ª–∏—è (Bougainvillia platygaster) –Ω–µ –≤—ã–±—Ä–∞—Å—ã–≤–∞–µ—Ç —è–π—Ü–∞ –≤ –≤–æ–¥—É. –Ø–π—Ü–∞ —Ä–∞–∑–≤–∏–≤–∞—é—Ç—Å—è —É –Ω–µ–µ –ø—Ä—è–º–æ –≤ –≥–æ–Ω–∞–¥–∞—Ö, –∑–¥–µ—Å—å –∂–µ –æ–±—Ä–∞–∑—É—é—Ç—Å—è –º–∞–ª–µ–Ω—å–∫–∏–µ –ø–æ–ª–∏–ø—ã, –∫–æ—Ç–æ—Ä—ã–µ –≤—ã–ø–æ—á–∫–æ–≤—ã–≤–∞—é—Ç –Ω–æ–≤—ã—Ö –º–∞–ª–µ–Ω—å–∫–∏—Ö –º–µ–¥—É–∑, –∞ –ø–æ—Å–ª–µ–¥–Ω–∏–µ –æ—Ç—à–Ω—É—Ä–æ–≤—ã–≤–∞—é—Ç—Å–º –∏ –ø–æ–∫–∏–¥–∞—é—Ç –ø–æ–ª–æ–≤—ã–µ –ø—É—Ç–∏ –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ—é –æ—Ä–≥–∞–Ω–∏–∑–º–∞. –°—Ä–µ–¥–∏ —Ä–∞—Å—Ç–µ–Ω–∏–π –ø—Ä–µ–æ–±–ª–∞–¥–∞—é—Ç –≤–∏–¥—ã, —É –∫–æ—Ç–æ—Ä—ã—Ö –≥–ª–∞–≤–Ω—ã–º –≤ –∂–∏–∑–Ω–µ–Ω–Ω–æ–º —Ü–∏–∫–ª–µ —è–≤–ª—è–µ—Ç—Å—è –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω—ã–π –∫–æ–º–ø–æ–Ω–µ–Ω—Ç, –Ω–æ –µ—Å—Ç—å –∏ –∏—Å–∫–ª—é—á–µ–Ω–∏—è, –Ω–∞–ø—Ä–∏–º–µ—Ä, —Ç—Ä–æ–ø–∏—á–µ—Å–∫–æ–µ —Ü–≤–µ—Ç–∫–æ–≤–æ–µ —Ä–∞—Å—Ç–µ–Ω–∏–µ-–ø–∞—Ä–∞–∑–∏—Ç –Ý–∞—Ñ—Ñ–ª–µ–∑–∏—è –ê—Ä–Ω–æ–ª—å–¥–∞, —É –∫–æ—Ç–æ—Ä–æ–≥–æ –ø–æ–±–µ–≥ (–≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–∞—è —Ñ–∞–∑–∞) –∏–º–µ–µ—Ç—Å—è —Ç–æ–ª—å–∫–æ –≤ —Ä–∞–Ω–Ω–µ–º –∑–∞—Ä–æ–¥—ã—à–µ. –ò–∑ –º–∞–ª–µ–Ω—å–∫–æ–≥–æ, —Å –º–∞–∫–æ–≤–æ–µ –∑–µ—Ä–Ω–æ, —Å–µ–º–µ–Ω–∏ —ç—Ç–æ–≥–æ —Ä–∞—Å—Ç–µ–Ω–∏—è –ø–æ—è–≤–ª—è–µ—Ç—Å—è —Å—Ä–∞–∑—É –º–∞–ª–µ–Ω—å–∫–∏–π —Ü–≤–µ—Ç–æ—á–Ω—ã–π –±—É—Ç–æ–Ω (–ø–æ–ª–æ–≤–∞—è —Ñ–∞–∑–∞) –±–µ–∑ —Å—Ç–µ–±–ª—è, –∫–æ—Ç–æ—Ä—ã–π —Ä–∞–∑–≤–∏–≤–∞–µ—Ç—Å—è –¥–µ–≤—è—Ç—å –º–µ—Å—è—Ü–µ–≤, –¥–æ—Å—Ç–∏–≥–∞–µ—Ç —Ä–∞–∑–º–µ—Ä–æ–≤ –±–æ–ª—å—à–æ–≥–æ –∫–æ—á–∞–Ω–∞ –∫–∞–ø—É—Å—Ç—ã –∏ —Ä–∞—Å–∫—Ä—ã–≤–∞–µ—Ç—Å—è –≤ –æ–≥—Ä–æ–º–Ω—ã–π —Ü–≤–µ—Ç–æ–∫, –¥–æ—Å—Ç–∏–≥–∞—é—â–∏–π 1 –∏ –≤ –¥–∏–∞–º–µ—Ç—Ä–µ –∏ –º–∞—Å—Å–æ–π –¥–æ 6 –∫–≥ [43].
–°—É—â–µ—Å—Ç–≤–µ–Ω–Ω–æ–µ —Å–Ω–∏–∂–µ–Ω–∏–µ —Ä–æ–ª–∏ –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π —Ñ–∞–∑—ã –ø—Ä–æ–∏–∑–æ—à–ª–æ –∏ –≤ –∂–∏–∑–Ω–µ–Ω–Ω–æ–º —Ü–∏–∫–ª–µ –ø–æ–∑–≤–æ–Ω–æ—á–Ω—ã—Ö –∂–∏–≤–æ—Ç–Ω—ã—Ö. –ï—Å–ª–∏ –Ω–µ–∫–æ—Ç–æ—Ä—ã–µ —Ñ–æ—Ä–º—ã –ø—Ä–∏–º–∏—Ç–∏–≤–Ω—ã—Ö —Ö–æ—Ä–¥–æ–≤—ã—Ö (—Ç—É–Ω–∏–∫–∞—Ç—ã - —Å–∞–ª—å–ø—ã –∏ –∞—Å—Ü–∏–¥–∏–∏) —Ä–∞–∑–º–Ω–æ–∂–∞—é—Ç—Å—è –≤ –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π —Ñ–∞–∑–µ –Ω–µ –º–µ–Ω–µ–µ —ç—Ñ—Ñ–µ–∫—Ç–∏–≤–Ω–æ, —á–µ–º –¥–∞–∂–µ –º–Ω–æ–≥–∏–µ –∫–∏—à–µ—á–Ω–æ–ø–æ–ª–æ—Å—Ç–Ω—ã–µ [27], —Ç–æ —É –ø–ª–∞—Ü–µ–Ω—Ç–∞—Ä–Ω—ã—Ö –º–ª–µ–∫–æ–ø–∏—Ç–∞—é—â–∏—Ö –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–∞—è —Ñ–∞–∑–∞ –ø—Ä–æ—è–≤–ª—è–µ—Ç —Å–µ–±—è –≤ —Ä–µ–¥–∫–æ–º —è–≤–ª–µ–Ω–∏–∏ —Ä–æ–∂–¥–µ–Ω–∏—è –æ–¥–Ω–æ—è–π—Ü–µ–≤—ã—Ö –±–ª–∏–∑–Ω–µ—Ü–æ–≤. –ù–µ–∫–æ—Ç–æ—Ä—ã–µ –≤–∏–¥—ã –±—Ä–æ–Ω–µ–Ω–æ—Å—Ü–µ–≤ (Dasipodini) –≤—Å–µ–≥–¥–∞ —Ä–æ–∂–¥–∞—é—Ç –ø–∞—Ä–Ω–æ–µ (–æ—Ç 2 –¥–æ 12) –∫–æ–ª–∏—á–µ—Å—Ç–≤–æ –æ–¥–Ω–æ—è–π—Ü–µ–≤—ã—Ö –±–ª–∏–∑–Ω–µ—Ü–æ–≤ [37, 44].
–í–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω—ã–π –æ—Ä–≥–∞–Ω–∏–∑–º –≤ —Ö–æ–¥–µ —ç–≤–æ–ª—é—Ü–∏–∏ —Å–Ω–∞—á–∞–ª–∞ –æ–∫–∞–∑–∞–ª—Å—è –ø–æ–º–µ—â–µ–Ω–Ω—ã–º –≤ –≥–æ–Ω–∞–¥—ã, –∑–∞—Ç–µ–º –±—ã–ª –∑–∞–∫–ª—é—á–µ–Ω –≤ —è–π—Ü–µ–≤—É—é –æ–±–æ–ª–æ—á–∫—É. –£ –ø–ª–∞—Ü–µ–Ω—Ç–∞—Ä–Ω—ã—Ö –º–ª–µ–∫–æ–ø–∏—Ç–∞—é—â–∏—Ö –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω—ã–π –æ—Ä–≥–∞–Ω–∏–∑–º - –±–ª–∞—Å—Ç–æ—Ü–∏—Å—Ç–∞ - –≤–Ω–æ–≤—å –æ–∫–∞–∑–∞–ª—Å—è –≤ –ø–æ–ª–æ–≤—ã—Ö –ø—É—Ç—è—Ö –º–∞—Ç–µ—Ä–∏–Ω—Å–∫–æ–≥–æ –æ—Ä–≥–∞–Ω–∏–∑–º–∞, –æ–±—Ä–∞–∑–æ–≤–∞–≤ –≤ –∑–æ–Ω–µ –∫–æ–Ω—Ç–∞–∫—Ç–∞ –ø–ª–∞—Ü–µ–Ω—Ç—É. –í–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–µ —Ä–∞–∑–º–Ω–æ–∂–µ–Ω–∏–µ –≤ —ç—Ç–æ–π —Ñ–∞–∑–µ —Å—Ç–∞–ª–æ —Å—Ä–∞–≤–Ω–∏—Ç–µ–ª—å–Ω–æ —Ä–µ–¥–∫–∏–º —è–≤–ª–µ–Ω–∏–µ–º, –Ω–æ –æ–Ω–æ –ø—Ä–æ–∏—Å—Ö–æ–¥–∏—Ç. –ü—Ä–∏—á–µ–º, —Å—É–¥—è –ø–æ —Ö–∞—Ä–∞–∫—Ç–µ—Ä—É —Å—Ä–∞—â–∏–≤–∞–Ω–∏—è –±–ª–∏–∑–Ω–µ—Ü–æ–≤—ã—Ö –ø–ª–∞—Ü–µ–Ω—Ç, –∞ —Ç–∞–∫–∂–µ –ø–æ—è–≤–ª–µ–Ω–∏—é –∏–Ω–æ–≥–¥–∞ —Ä–∞–∑–ª–∏—á–Ω—ã—Ö —Å–∏–∞–º—Å–∫–∏—Ö –±–ª–∏–∑–Ω–µ—Ü–æ–≤ [45], —ç—Ç–æ —Ä–∞–∑–º–Ω–æ–∂–µ–Ω–∏–µ –ø—Ä–æ–∏—Å—Ö–æ–¥–∏—Ç –æ—Ç–Ω—é–¥—å –Ω–µ —Ç–æ–ª—å–∫–æ –Ω–∞ —Å—Ç–∞–¥–∏–∏ –¥–≤—É—Ö –±–ª–∞—Å—Ç–æ–º–µ—Ä–æ–≤, –∫–∞–∫ –ø—Ä–∏–Ω—è—Ç–æ —Å—á–∏—Ç–∞—Ç—å, –Ω–æ –∏ –Ω–∞ –±–æ–ª–µ–µ –ø–æ–∑–¥–Ω–∏—Ö —Å—Ç–∞–¥–∏—è—Ö —Ä–∞–∑–≤–∏—Ç–∏—è –æ—Ä–≥–∞–Ω–∏–∑–º–∞ –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π —Ñ–∞–∑—ã, –∫–∞–∫ –Ω–∞–ø—Ä–∏–º–µ—Ä —É –±—Ä–æ–Ω–µ–Ω–æ—Å—Ü–µ–≤ [37].
–ü–æ –Ω–∞—à–µ–º—É –º–Ω–µ–Ω–∏—é, —Ñ–∞–∫—Ç —Å—É—â–µ—Å—Ç–≤–æ–≤–∞–Ω–∏—è –≤–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–æ–π –∏ –ø–æ–ª–æ–≤–æ–π —Ñ–∞–∑ –≤ –∂–∏–∑–Ω–µ–Ω–Ω–æ–º —Ü–∏–∫–ª–µ –∂–∏–≤–æ—Ç–Ω—ã—Ö —è–≤–ª—è–µ—Ç—Å—è –æ—Å–Ω–æ–≤–æ–π –¥–ª—è —Ñ–æ—Ä–º–∏—Ä–æ–≤–∞–Ω–∏—è –ø—Ä–µ–¥—Å—Ç–∞–≤–ª–µ–Ω–∏–π –æ –±–∏–æ–ª–æ–≥–∏—á–µ—Å–∫–æ–π —Å—É—â–Ω–æ—Å—Ç–∏ —Ñ–µ–Ω–æ–º–µ–Ω–∞ –∑–ª–æ–∫–∞—á–µ—Å—Ç–≤–µ–Ω–Ω–æ–≥–æ —Ä–æ—Å—Ç–∞.
–í–µ–≥–µ—Ç–∞—Ç–∏–≤–Ω–∞—è –∏ –ø–æ–ª–æ–≤–∞—è —Ñ–∞–∑—ã –≤ –∂–∏–∑–Ω–µ–Ω–Ω–æ–º —Ü–∏–∫–ª–µ –º–Ω–æ–≥–æ–∫–ª–µ—Ç–æ—á–Ω—ã—Ö –æ—Ä–≥–∞–Ω–∏–∑–º–æ–≤
–í –∫–Ω. –ë–∏–æ–ª–æ–≥–∏—è –º–∞—Ä–∫–µ—Ä–æ–≤ —Ä–∞–∫–∞ –∏ –±–µ—Ä–µ–º–µ–Ω–Ω–æ—Å—Ç–∏. –ö–∏–µ–≤ : –ù–∞—É–∫. –î—É–º–∫–∞, 1990.252 —Å. ISBN 5-12-001790-8. —Å. 14-19
–í.–ë.–í–∏–Ω–Ω–∏—Ü–∫–∏–π, –õ.–ê.–ü–æ–ª–∏—â—É–∫